
 Download:
Download:




HTML
-
Introduction: Bacillus Calmette–Guérin (BCG) is the only approved tuberculosis (TB) vaccine; however, it has partial efficacy and time-limited protection. Subunit booster vaccines may enhance the effects of BCG; however, their efficacy depends on their composition.
Methods: Six-week-old BALB/c mice received primary immunization with BCG, followed by EPCP009 protein boosters (mixed with DDA/poly:IC [DP] adjuvant) once, twice, or thrice (2-week intervals). Six weeks after the first immunization, blood and spleen samples were collected. Levels of antibodies and cytokines were assessed via enzyme-linked immunosorbent assay (ELISA), Luminex, and enzyme-linked immunospot (ELISpot) The ability of splenocytes to inhibit Mycobacterium tuberculosis (Mtb) was assessed by MGIA, and pathological analyses were conducted on the liver, spleen, and lung. In addition, mice were immunized with EPCP009 or H4 protein separately. BCG was administered intravenously 6 weeks later, and spleens were collected at 3.5 weeks to quantify the BCG bacterial load.
Results: BCG combined with a single EPCP009 dose provided better short-term (6-week) and long-term (12-week) protection than multiple doses. During the long-term process, the levels of purified protein derivative (PPD)-specific interferon (IFN)-γ and granulocyte-macrophage colony-stimulating factor (GM-CSF) remained relatively high. No significant differences in the BCG colony counts were observed among the EPCP009/DP, phosphate-buffered saline (PBS), and DP groups.
Conclusions: These findings demonstrate that BCG vaccination combined with a single dose of EPCP009 enhances protective efficacy against TB. This study provides a critical theoretical basis for designing booster immunization strategies for TB vaccines.
-
Tuberculosis (TB) is the second leading cause of death from a single infectious agent, following coronavirus disease 2019 (COVID-19) (1). According to the World Health Organization (WHO) Global TB Report 2023, more than 10.6 million individuals, with approximately 90% of them adults, contracted TB by 2022 (2). A quarter of the world’s population is estimated to be infected with Mycobacterium tuberculosis (Mtb). The COVID-19 pandemic has reduced healthcare accessibility for patients with TB, setting back global TB prevention and control efforts (3). Developing new TB vaccine strategies is essential for achieving the ambitious goal of ending TB by 2030.
Mycobacterium bovis Bacillus Calmette–Guérin (BCG) is the only approved preventive vaccine offering protection against severe TB for young children. However, its efficacy in adults remains unclear. The WHO does not recommend BCG revaccination in adults because of insufficient evidence supporting its effectiveness (4). Numerous TB vaccines are currently under development, with 18 undergoing clinical trials worldwide. Among these, the subunit TB vaccine M72/AS01E has demonstrated 49.7% efficacy [90% confidence interval (CI), 12.1, 71.2] against active pulmonary TB in adults; however, its efficacy requires further evaluation (5). Clinical trials of the H4:IC31 and M72/AS01E adjuvant protein subunit vaccines co-administered with BCG show promising results and offer prospects for different immunization strategies. Viral vector and mRNA vaccines have received special attention as emerging platforms. The ChAdOx1 85A viral vector vaccine induced Ag85-specific CD4+T and CD8+T cells and was well tolerated and immunogenic in a phase I clinical trial (6). Animal models have demonstrated both preventive and therapeutic effects of mRNA vaccines in zebrafish. However, the clinical protective effect of the existing single-dose virus vector vaccine is not ideal, and the mRNA vaccine still faces challenges, such as stability.
Unlike attenuated or live vaccines, subunit protein vaccines have clear antigenic components and high safety profiles, making them particularly suitable for HIV-infected and immunocompromised populations. However, subunit vaccines often require multiple booster doses to maximize efficacy, posing challenges in TB-endemic regions with limited resources. Heterologous prime-boost regimens using BCG and different antigen-expressing vectors can induce more robust T-cell and humoral responses (7). With global BCG coverage reaching 90%, new TB vaccine development focuses on designing booster vaccines following initial BCG immunization. Most TB subunit vaccines in clinical trials, such as H4/IC31 and H56/IC31, are efficacious when used as boosters after BCG vaccination (8–9). However, no approved immune-enhancing vaccines are currently available for use with BCG; the reasons remain unclear. Most subunit vaccines share some or all antigenic components with BCG, which may inhibit BCG growth and immune responses.
As part of our TB vaccine development program, we designed the EPCP009 fusion protein, which is composed of four antigens that are either not expressed or expressed at low levels in BCG. This design broadens the range of immune protection without interfering with BCG-mediated immune responses. We previously demonstrated superior effectiveness of BCG prime-EPCP009 booster regimen over the rBCG-EPCP009 regimen (10). Herein, BALB/c mice were used to investigate the immunogenicity, efficacy, and safety of BCG in combination with single, double, and triple doses of EPCP009/dimethyl-dioctyldecylammonium bromide/polyinosinic-polycytidylic acid (DDA/poly:IC). In addition, we assessed whether EPCP009 inhibited BCG growth, elucidating its potential as a BCG booster vaccine.
Mycobacterium bovis BCG-China and Mycobacterium tuberculosis H37Rv (ATCC27294) strains were maintained at −80 °C and cultured on Difco™ Middlebrook 7H9/7H10 agar (BD, USA) supplemented with 10% OADC, 0.5% glycerol, and 0.05% tween 80. Recombinant pET28a(+) plasmid expressing H4 antigen (TB10.4-Ag85B fusion protein) was constructed in our laboratory and transformed into Escherichia coli DH5α/BL21(DE3).
Female BALB/c mice (6 weeks old, specific-pathogen-free) were purchased from Beijing HFK Bioscience Co., Ltd. (Beijing, China). Mice were acclimatized for 5 days in pathogen-free cages before random allocation to five groups (n=12/group).
Each vaccine dose containing 10 μg of EPCP009 protein was formulated in phosphate-buffered saline (PBS) (pH 7.4) with 250 μg of DDA/50 μg of poly:IC adjuvant. BCG-China [1×106 colony-forming units (CFU)] or EPCP009 formulations were administered intradermally (100 μL) or subcutaneously (200 μL), respectively. Booster immunizations with EPCP009 were administered at 2-week intervals after BCG priming (BCG+1×, BCG+2×, or BCG+3×EPCP009 groups) (11). The control group received the DDA/poly:IC (Sigma, Germany) adjuvant or PBS alone (Figure 1).
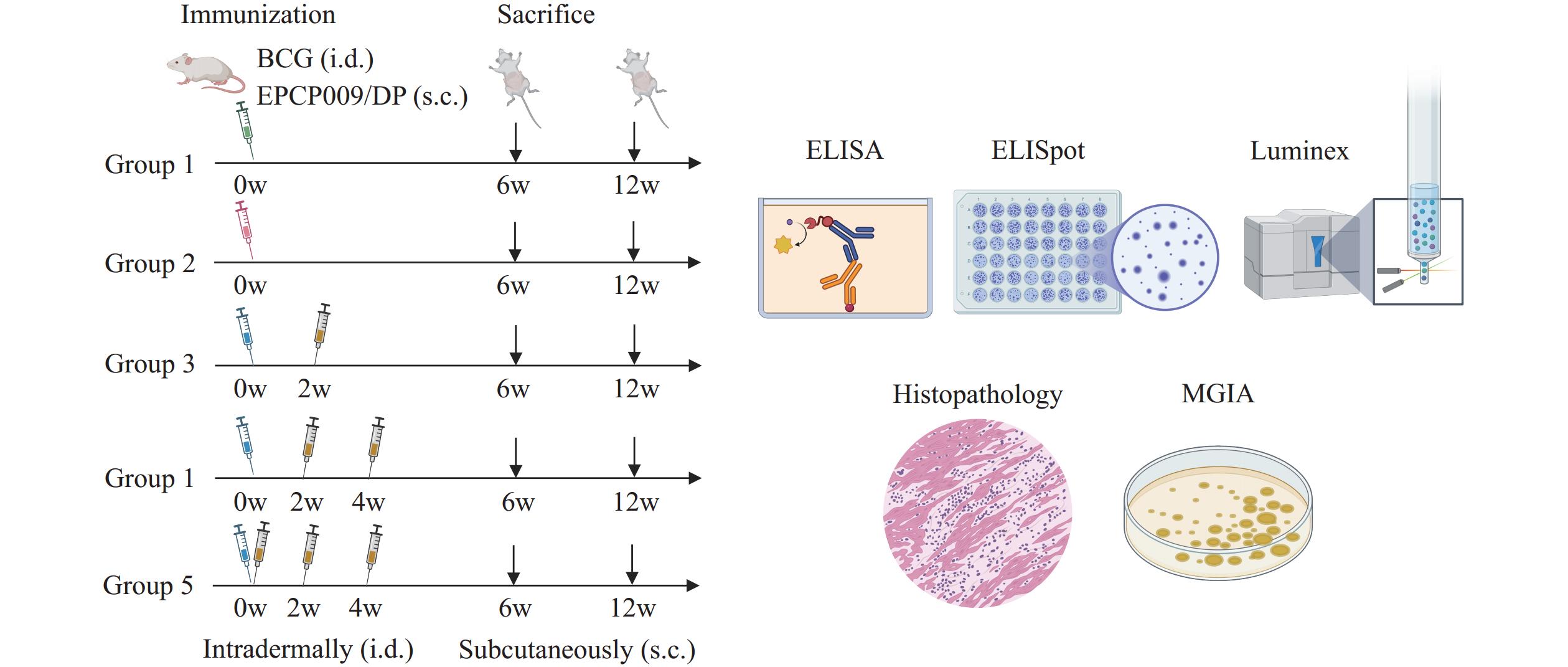 Figure 1.
Figure 1.Vaccination and testing regimen schedules.
Abbreviation: ELISA=enzyme-linked immunosorbent assay; ELISPOT=enzyme-linked immunospot; MGIA=mycobacterial growth inhibition assay.Spleens were harvested at 6/12 weeks post-immunization, minced through 70-μm strainers (BD, USA), and centrifuged (800 ×g, 30 min). The cells were washed twice with RPMI 1640 (Gibco, USA), resuspended in 10% fetal bovine serum-RPMI, and adjusted to 2×105 cells/well (ELISpot/Luminex) or 5×106 cells/well (MGIA).
ELISpot: Splenocytes (2×105/well) were stimulated with 2 μg of PPD for 20 h (37 °C, 5% CO2) and detected using the ELISpot assay kit (Dakewe, China) (12).
Luminex: Supernatants from PPD-stimulated splenocytes (10 μg/well, 24 h) were analyzed for nine cytokines (interferon [IFN]-γ, interleukin [IL]-2, tumor necrosis factor [TNF]-α, IL-17, granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-12, IL-4, IL-6, IL-10) using the magnetic bead assay (R&D Systems, USA).
Enzyme-linked immunosorbent assay (ELISA): A 96-well ELISA plate was coated with 200 ng of EPCP009 antigen, horseradish peroxidase-labeled goat anti-mouse IgG was used as the secondary antibody, and 3,3',5,5'-tetramethylbenzidine substrate was added to develop color.
Spleen, liver, and lung tissues collected at 6 weeks of age were fixed in 4% paraformaldehyde for hematoxylin and eosin (H&E) staining ( Beijing Kaike Green Technology Co., Ltd., China).
Splenocytes (5×106 cells) were co-cultured with Mtb H37Rv (50 CFU) for 72 h at 37 °C and 5% CO2. After incubation, the supernatant was removed by centrifugation at 4,000 rpm for 10 min. The cells were lysed by adding 500 μL of pre-cooled sterile water in an ice bath per well to release intracellular bacteria, and 50 μL was spread on the 7H10 medium containing 10% OADC. Six replicates were set up for each sample.
The H4 protein (containing the Mtb/BCG-shared antigens TB10.4 and Ag85B) served as a positive control in assessing cross-immunogenicity between EPCP009 and BCG (8). Recombinant EPCP009 and H4 were expressed in E. coli BL21(DE3) cells using pET28a(+) vectors with kanamycin selection (25 μg/mL). Protein induction was performed with 1 mmol/L of IPTG (Sigma, Germany) for 4.5 h at 37 °C, followed by nickel-affinity chromatography purification (GE Healthcare, USA) from inclusion bodies.
Mice were divided into four groups: PBS, DDA/poly:IC (DP) adjuvant (250 μg DDA/50 μg poly:IC), EPCP009/DP (10 μg protein + adjuvant), and H4/DP (10 μg protein + adjuvant). Immunization was administered subcutaneously at 0-, 2-, and 4-week intervals. Six weeks after immunization, BCG-China (1×107 CFU) was administered intravenously. The spleens were collected at 3.5 weeks, lysed, and the BCG bacterial load was quantified using 7H10 medium.
Data are presented as mean ± standard deviation (GraphPad Prism 8.0, San Diego, California, USA, www.graphpad.com). Group comparisons were performed using one-way analysis of variance (ANOVA) with Tukey’s multiple comparison test.
The amounts of IFN-γ (A,C) and IL-4 (B,D) secreted by mouse splenic lymphocytes at 6 and 12 weeks were determined using the ELISpot assay.
Six and 12 weeks after the initial immunization, mouse splenic lymphocytes were stimulated with PPD (Figure 2). All vaccine groups produced higher levels of the protective cytokine IFN-γ at both 6 (P<0.05) and 12 (P<0.05) weeks than the PBS group. The BCG+1×EPCP009 group induced higher IFN-γ production than the BCG (P<0.001) and BCG+2×EPCP009 (P<0.05) groups at both time points. The BCG+1×EPCP009 group induced comparable production of IFN-γ to the BCG+3×EPCP009 group at 6 weeks (P=0.041), and significantly higher than the BCG+3×EPCP009 group at 12 weeks (P<0.001). IL-4 levels in the BCG+1×EPCP009 group were significantly higher than those in the PBS (P<0.001) and BCG (P<0.001) groups at 6 weeks, but comparable to those in the BCG+2×EPCP009 (P=0.919) and BCG+3×EPCP009 (P=0.058) groups. IL-4 levels in the BCG+1×EPCP009 group were comparable to those in the BCG (P=0.953) and BCG+2×EPCP009 (P=0.970) groups at 12 weeks but were significantly higher than those in the BCG+3×EPCP009 group (P=0.018).
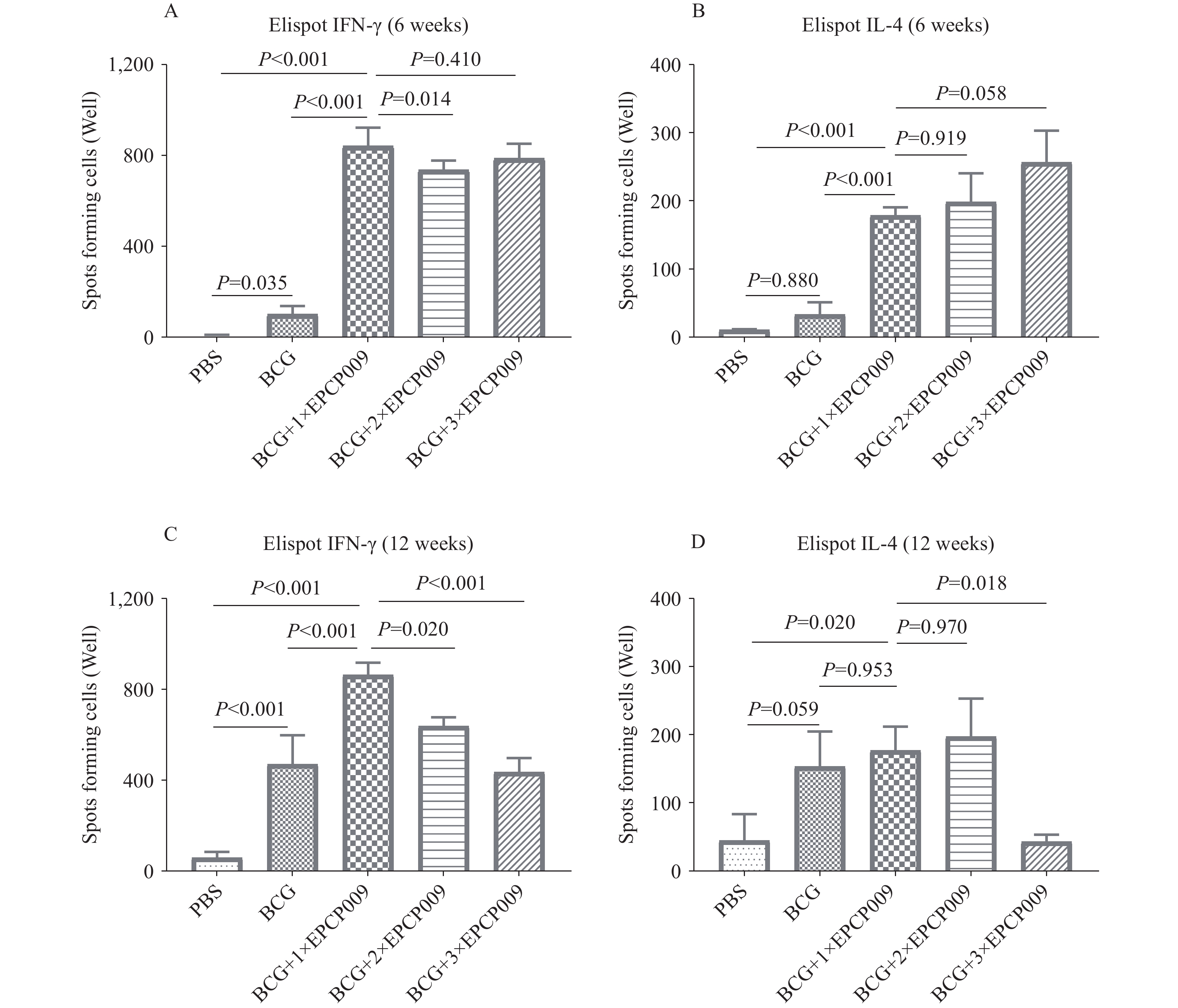 Figure 2.
Figure 2.PPD-specific IFN-γ and IL-4 secreted by mice splenic lymphocytes. (A) IFN-γ and IL-4 (B) secreted at 6 weeks; (C) IFN-γ and IL-4 (D) secreted at 12 weeks.
Abbreviation: PPD=purified protein derivative; ELISpot=enzyme-linked immunospot.Six weeks post-immunization, mouse splenic lymphocytes were stimulated with PPD (Figure 3 and
Supplementary Figure S1 ). The BCG+1×EPCP009 group produced higher levels of IFN-γ, IL-2, TNF-α, IL-17, GM-CSF, IL-12, IL-4, IL-6, and IL-10 than the PBS and BCG groups (both P<0.05). In addition, the BCG+1×EPCP009 group induced higher levels of IFN-γ, IL-2, IL-12, IL-17, and GM-CSF than the BCG+2×EPCP009 group (P<0.05). Compared with the BCG+3×EPCP009 group, the BCG+1×EPCP009 group had significantly higher IL-2 levels (P<0.001) but lower IL-6 levels (P<0.001).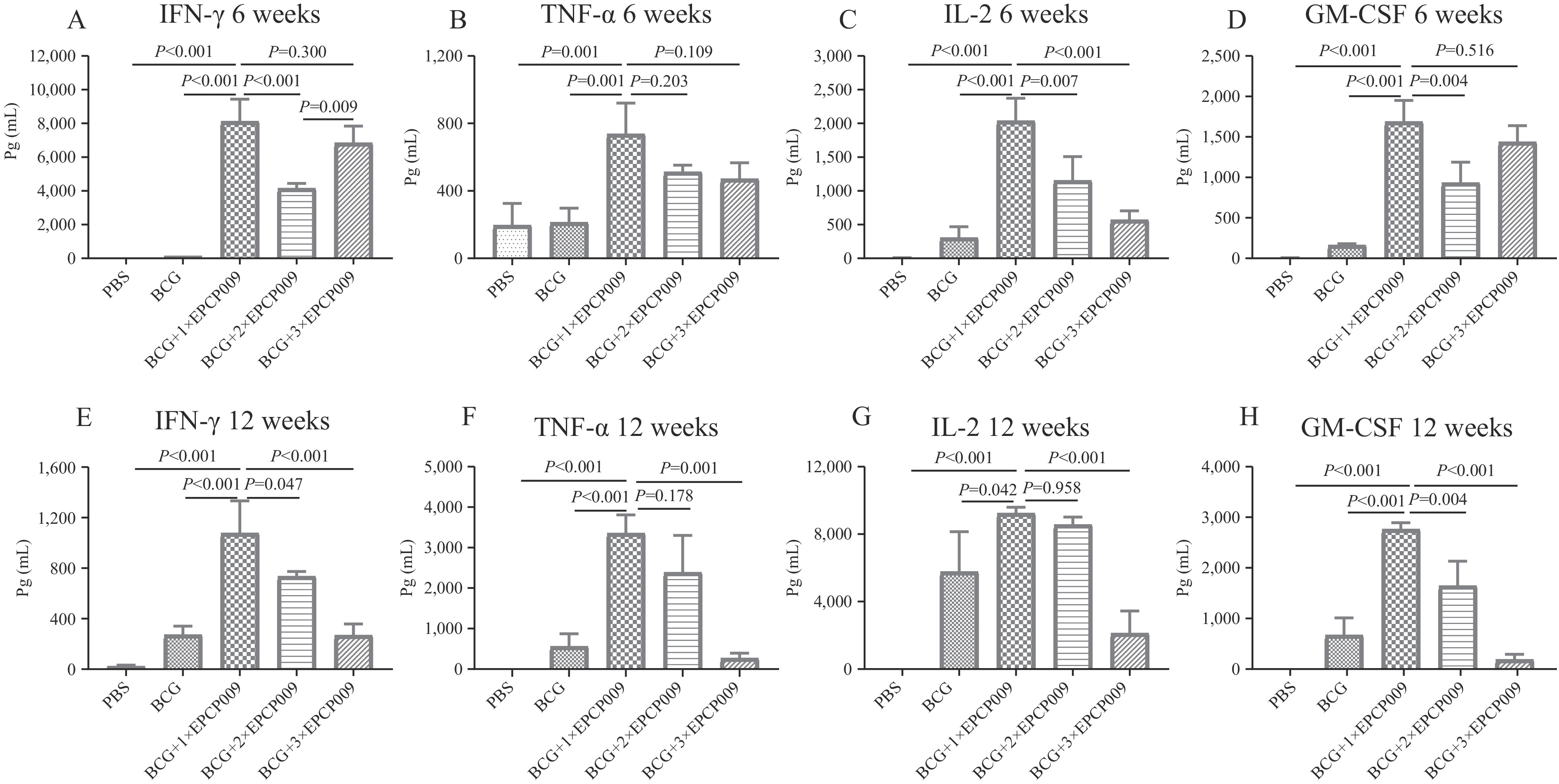 Figure 3.
Figure 3.Levels of cytokines secreted by PPD-stimulated spleen lymphocytes in mice 6 and 12 weeks from initial immunization. (A) IFN-γ, (B) TNF-α, (C) IL-2 and (D) GM-CSF secreted at 6 weeks; (E) IFN-γ, (F) TNF-α, (G) IL-2 and (H) GM-CSF secreted at 12 weeks.
At 12 weeks post-immunization, the BCG+1×EPCP009 group induced higher levels of IFN-γ, GM-CSF, and IL-4 than the PBS, BCG, BCG+2×EPCP009, and BCG+3×EPCP009 groups (P<0.05) (Figure 3 and
Supplementary Figure S1 ). Furthermore, the BCG+1×EPCP009 group produced higher levels of IL-17 than the BCG+2×EPCP009 and BCG+3×EPCP009 groups (P<0.05).The ability and duration of immune-mediated inhibition of Mtb growth in vitro were evaluated, for which splenocytes were isolated from immunized mice and co-cultured with H37Rv at 6 and 12 weeks following initial immunization. The bacterial colonies were quantified and expressed as log10 colony-forming units (lgCFUs) per sample. At 6 weeks post-immunization, the BCG+1×EPCP009 group displayed significantly lower colony counts than the PBS, BCG, and BCG+2×EPCP009 groups (P<0.05), and slightly lower colony counts than the BCG+3×EPCP009 group (P=0.063). At 12 weeks, the BCG+1×EPCP009 group had significantly lower colony counts than the other groups (P<0.001) (Figure 4).
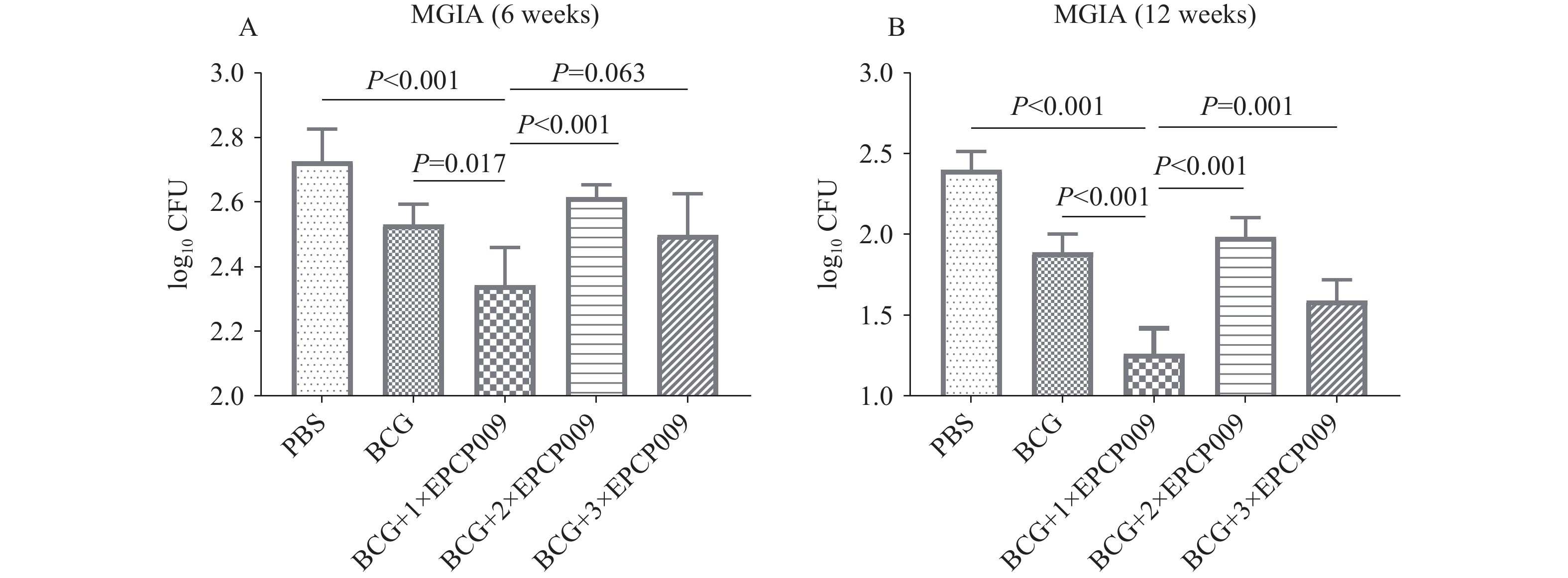 Figure 4.
Figure 4.In vitro growth inhibition of Mtb H37Rv with splenocytes from immunized BALB/c mice at (A) 6 weeks and (B) 12 weeks.
Abbreviation: MGIA=mycobacterial growth inhibition assay.At 6 weeks post-immunization, histological analysis of the liver, spleen, and lung tissues revealed no significant between-group differences. All organs exhibited normal morphology with no evidence of cellular edema or necrosis (Figure 5).
 Figure 5.
Figure 5.Safety analysis of BCG prime-EPCP009 booster with different number of immunizations. After 6 weeks of the last immunization, the liver (A, B, C, D, E), spleen (F, G, H, I, J), and lungs (K, L, M, N, O) of the PBS, BCG, BCG+1×EPCP009, BCG+2×EPCP009, BCG+3×EPCP009 groups were aseptically isolated and stained with H&E.
Abbreviation: H&E=hematoxylin and eosin.The H4/DP group demonstrated significantly fewer BCG colonies in the spleen than the PBS (P=0.005) and DP (P=0.004) groups, indicating cross-immunity between H4 and BCG (Figure 6). In contrast, the EPCP009/DP group displayed no significant difference in BCG colony counts compared to the PBS (P=0.967) and DP (P=0.986) groups, indicating that EPCP009 did not inhibit BCG growth or colonization.
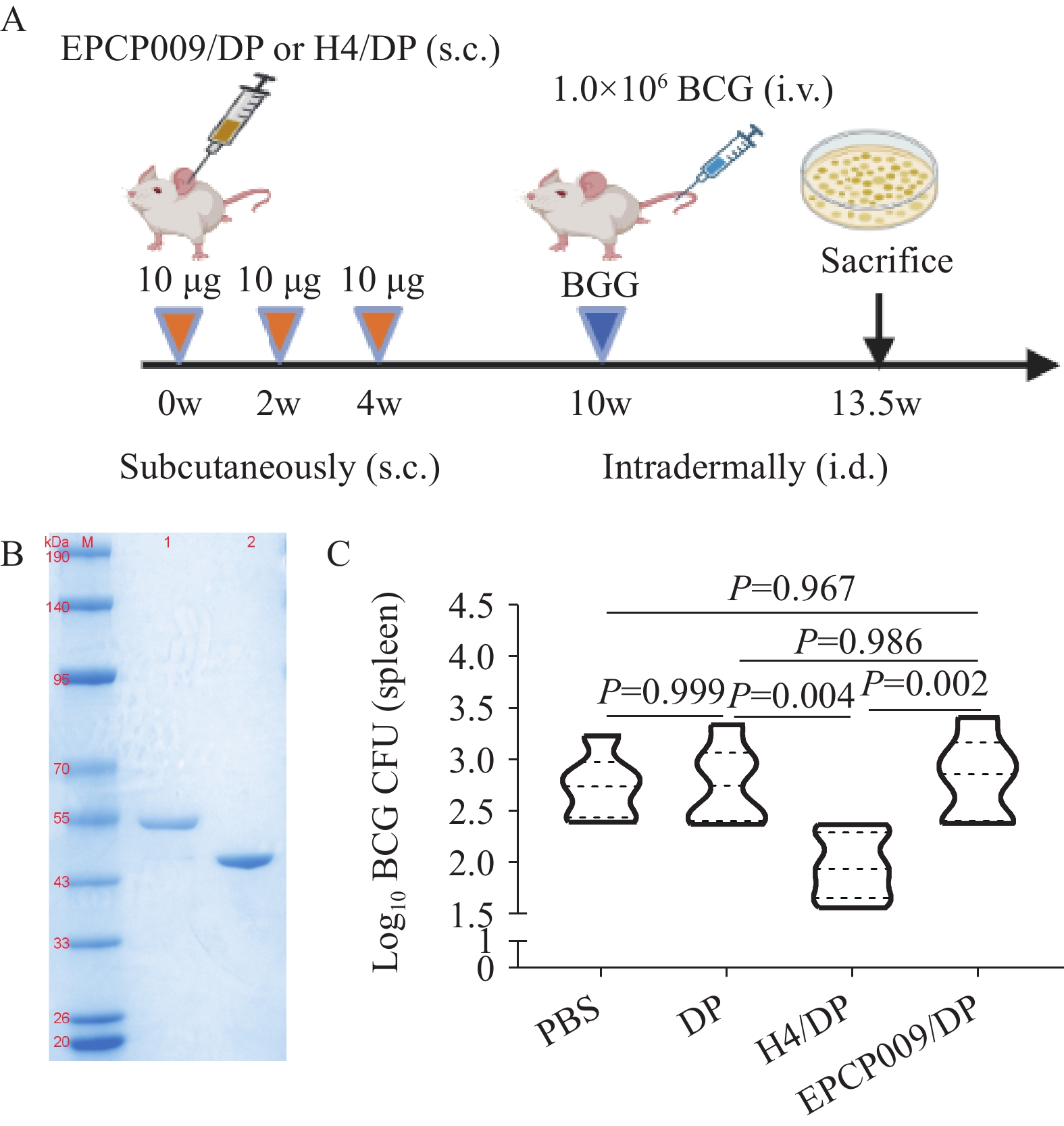 Figure 6.
Figure 6.Verification of cross-immune responses between EPCP009 and H4 proteins and BCG. (A) Cross immunization schemes. (B) Protein expression of EPCP009 and H4. (C) The number of BCG colonies in the spleens of BCG-inoculated mice after immunization with PBS, DP, H4/DP and EPCP009/DP.
Note: For (B), lane M, molecular weight marker of the pre-stained protein; lane 1, purified EPCP009 protein, 51 kD; lane 2, purified H4 protein, 46 kD.
-
After decades of global efforts, more than a dozen TB vaccines have reached clinical trials, primarily falling into two categories: whole mycobacterium-derived vaccines (live attenuated and inactivated) and subunit vaccines (13). Subunit vaccines, which are particularly promising BCG boosters, enhance and prolong BCG-induced immunity. Notably, subunit vaccines such as H4-IC31, Mtb72F/AS02A, and ID93/GLA-SE offer superior protection compared with BCG alone (14). A limited number of antigens can be selected for subunit vaccines; therefore, antigen selection and immunization frequencies are critical. EPCP009, introduced herein, was administered once after BCG vaccination to significantly improve immunization efficacy.
EPCP009 uniquely combines four antigens, CFP-10, ESAT-6, PPE18, and PstS1, none of which is expressed or minimally expressed in BCG. This broad antigen spectrum, including the highly immunogenic PstS1, which is not yet featured in clinical TB vaccines, enhances the immune response breadth (15). Unlike certain subunit vaccines that risk immune-mediated side effects due to antigens shared with BCG, EPCP009 avoids cross-reactivity, preserving BCG colonization and efficacy. Comparative studies with the H4 vaccine, which inhibits BCG growth, confirmed the absence of EPCP009 interference with BCG, highlighting its suitability as a BCG booster.
Given the intracellular nature of the Mtb, cellular immunity is paramount (16). EPCP009, in a single dose post-BCG, significantly elevated protective cytokines (IFN-γ, GM-CSF, and IL-2) at both 6 and 12 weeks, outperforming multiple-dose regimens and BCG alone. Thus, a durable Th1-biased immune response is crucial for Mtb control. Moreover, the MGIA assay revealed that a single booster immunization with EPCP009 after BCG immunization inhibited Mtb growth in vitro, and this inhibitory effect remained significant, especially after 12 weeks of immunization. The immunized groups produced higher antibody levels for longer durations (6 and 12 weeks) (
Supplementary Figure S2 ). In addition, BCG primary immunization with EPCP009 multiple booster immunization significantly decreased protective cytokines such as IFN-γ, TNF-α, IL-2, and GM-CSF, and other protective cytokines at week 12, with poor protection, which may be attributed to immune tolerance caused by multiple booster immunization. In conclusion, booster immunization with EPCP009 after BCG primary immunization has the potential for long-term protection.Despite its promise, this study acknowledges its limitations, including the need for validation across different BCG strains and animal models to better predict clinical efficacy. Nonetheless, EPCP009’s single-dose efficacy, both short- and long-term, makes it a highly promising candidate for an effective TB vaccine strategy, offering a significant advancement over current options.
-
All animal experiments were conducted in accordance with the animal welfare guidelines and approved by the Animal Experimental Ethical Committee of the Chinese Center for Disease Control and Prevention (2022-CCDC-IAC-026).
| Citation: |

|