
 Download:
Download:




-
Salmonella spp. infections represent a significant global public health concern, with non-typhoidal Salmonella (NTS) being the leading cause of diarrhea in China (1). Among NTS strains, Salmonella 4,[5],12:i:-, a monophasic variant of S. Typhimurium, has garnered considerable attention due to its rapid global dissemination over the past two decades and its substantial impact on human health (2−4). In Europe, Salmonella 4,[5],12:i:- has emerged as the third most common cause of non-typhoidal Salmonella infections since 2013 (2,5). In the United States, it ranks among the top five serotypes associated with salmonellosis (6). Data from the Chinese Pathogen Identification Network (2001–2022) revealed that Salmonella 4,[5],12:i:- is the third most prevalent serotype among NTS strains isolated from human sources in China (7).
The overall resistance rate of Salmonella in China during the past five decades across animal, food, and human sectors has reached 73.63%, with higher resistance levels to ciprofloxacin and cefotaxime observed domestically compared to international reports (8). Resistance to fluoroquinolones and third-generation cephalosporins in Salmonella was designated a high-priority issue by the WHO in 2017 and 2024 (9−10), underscoring the global burden of antimicrobial resistance (AMR) and the need for comprehensive studies on its transmission, treatment, and prevention. Previous research has demonstrated that Salmonella 4,[5],12:i:- is predominantly multidrug resistant, characterized by the ASSuT resistance profile and ST34 genotype (11). Evolutionary studies suggest that multidrug-resistant Salmonella 4,[5],12:i:- ST34 in the United States originated from multiple introductions from Europe (12), whereas in Australia, two lineages of Salmonella 4,[5],12:i:- are travel-associated while the third is locally endemic (13). Understanding the resistance patterns, evolutionary changes, and potential transmission dynamics of Salmonella 4,[5],12:i:- is critical for controlling resistance and preventing outbreaks. To date, however, few nationwide reports have examined the antimicrobial resistance and genomic characteristics of Salmonella 4,[5],12:i:- from human sources in China. Accordingly, to explore the genomic and phenotypic characteristics of Salmonella 4,[5],12:i:-, we conducted a retrospective analysis of resistance phenotypes, resistance determinants, and genomic heterogeneity of 2,736 Salmonella 4,[5],12:i:- isolates from diarrheal cases in China from 2017 to 2023. Our findings revealed that Salmonella 4,[5],12:i:- exhibits high genetic diversity and is widely dispersed throughout China, with high levels of resistance to multiple key antimicrobials, an increasing resistance trend over the years, and an upsurge in carbapenem resistance. These results provide valuable insights into the spatiotemporal distribution, antimicrobial resistance, genetic evolution, and epidemic patterns of Salmonella 4,[5],12:i:- in China.
-
All data, including geographical information, antimicrobial susceptibility testing results, and whole-genome sequences, were obtained from the National Pathogen Identification Network (CPIN) database.
Based on an active national hospital-based surveillance system, a total of 2,736 Salmonella 4,[5],12:i:- strains were isolated from diarrheal patients across 29 provinces in China between 2017 and 2023 (
Supplementary Figure S1 ). Most isolates were obtained from Zhejiang and Guangdong Provinces during the past two years. Young children constituted the predominant demographic group (≤5 years, 1,806 isolates; 6–14 years, 125 isolates; 15–64 years, 500 isolates; ≥65 years, 232 isolates). -
Antimicrobial susceptibility testing was conducted using the broth microdilution method for 17 agents across 11 classes, including ciprofloxacin (CIP), cefotaxime (CTX), meropenem (MEM), and polymyxin E (CT). Results were interpreted as susceptible, intermediate, or resistant using modified breakpoints (
Supplementary Table S1 ) according to the Clinical and Laboratory Standards Institute (CLSI) guidelines. Multidrug resistance (MDR) was defined as resistance to ≥3 classes of antimicrobials. -
The assembled genomic sequences from CPIN underwent quality control, resulting in a final dataset of 2,736 genomes. A maximum likelihood tree based on single nucleotide polymorphisms (SNPs) in the core genome was constructed using Parsnp software (14). The tree was visualized and modified using iTOL (https://itol.embl.de). PubMLST (https://pubmlst.org/) and ResFinder (https://genepi.food.dtu.dk/resfinder) were used for multilocus sequence typing (MLST) and resistance gene identification, respectively. Plasmid comparisons were conducted via NCBI BLASTn and visualized with Easyfig.
-
Statistical analysis was performed using SAS (Version 9.2; SAS Institute, Cary, NC, USA). The Chi-square test or Fisher’s exact test was employed to compare resistance rates, with a significance level of α=0.05. Trends in resistance rates were analyzed using the Cochran‒Armitage trend test. Kappa statistics were used to assess the concordance between resistance genes and phenotypes.
-
A total of 2,736 Salmonella 4,[5],12:i:- isolates demonstrated varying degrees of resistance to 17 antibiotics (
Supplementary Table S2 ). The overall resistance rate was 98.83%, with the highest resistance observed for streptomycin (91.91%), followed by tetracycline (90.42%) and ampicillin (86.77%). Resistance to cefotaxime reached 25.74% and ciprofloxacin 11.36%, while azithromycin resistance was 8.26%, indicating reduced sensitivity to this key macrolide. Low resistance rates (<5.00%) were observed for polymyxin E, ertapenem, meropenem, ceftazidime, and amikacin. Notably, we identified 15 meropenem-resistant strains, predominantly from infants aged ≤5 years in 2023 (10/15), which remained susceptible to colistin, azithromycin, tigecycline, and amikacin. The majority of these strains originated from Guangdong province (5/15), with four isolated in 2023 and one in 2022. The remaining 10 strains were distributed across 8 provinces in different years (two from Guangxi in 2023, two from Sichuan in 2023, and one each from Beijing 2023, Fujian 2022, Henan 2023, Jilin 2020, Jiangsu 2023, and Yunnan 2023, respectively). From 2020 to 2023, Salmonella 4,[5],12:i:- maintained high resistance levels to three antibiotics (ampicillin, streptomycin, and tetracycline;Supplementary Figure S2 ) and exhibited significant increasing trends in resistance to seven antibiotics (AMP, CHL, SXT, MEM, AZM, STR, AMS) over time (Z values: –1.9706, –6.2794, –4.8912, –1.9336, –3.438, –1.8369, and –1.7365; all P<0.05;Supplementary Figure S2 ).The MDR rate was 87.98% (2,407/2,736), with one isolate exhibiting resistance to as many as 15 antibiotics (CHL-SXT-CT-ETP-CTX-CTZ-TET-TIG-CIP-NAL-AZM-AMI-STR-AMP-AMS). The dominant resistance profile was AMP-STR-TET (AST, 16.30%, 446/2,736;
Supplementary Table S3 ). No significant differences in overall resistance rate or MDR rate were found across different age groups (both exceeding 98.00% and 85.00%, respectively); however, resistance rates to chloramphenicol, cefotaxime, and ceftazidime differed significantly (χ2=11.83, 23.414, and 20.054, respectively; all P<0.05). Compared with adults, infants exhibited significantly higher resistance to chloramphenicol, cefotaxime, and ceftazidime (50.17% vs. 41.97%, 28.52% vs. 19%, and 14.68% vs. 8.02%, respectively; all P<0.05). Additionally, significant differences in resistance rates were observed among provinces (P<0.05), with Sichuan Province showing the highest resistance rate (100%), followed by Guangdong Province (99.77%), while Jiangxi Province exhibited the lowest resistance rate (97.45%). These findings suggest that resistance rates may increase with the level of economic development, consistent with previous reports (15–16). Beyond economic factors, antibiotic usage practices, hygiene conditions, and climate are considered contributors to antimicrobial resistance development and variations in Salmonella across different geographic areas and time periods, as well as in other Enterobacterales (17–19). -
Most (96.49%) Salmonella 4,[5],12:i:- isolates carried resistance genes encompassing 11 classes of antibiotics (a total of 112 different resistance genes, excluding aac(6')-Iaa), with the highest prevalence observed for β-lactam genes (85.16%), tet(B) (75.15%), and sul2 (73.50%). Statistical analysis revealed that the prevalence of 67 resistance genes, primarily associated with aminoglycosides, beta-lactams, and macrolides, increased significantly annually (Z values of −2.024, −4.1929, and −2.4495, respectively; all P<0.05; Figure 1). The mcr-1 and mcr-9 genes, conferring resistance to colistin, were identified in 13 isolates collected between 2020 and 2023, predominantly from infants (9/13). All mcr-1-positive Salmonella 4,[5],12:i:- isolates harbored 4–6 classes of AMR genes and belonged to ST34, with most exhibiting MDR phenotypes, including resistance to colistin, ampicillin, chloramphenicol, streptomycin, tetracycline, trimethoprim-sulfamethoxazole, and cefotaxime. High concordance was observed between resistance phenotypes and resistance genes for chloramphenicol, beta-lactams, and quinolones (kappa >0.6) (
Supplementary Table S4 ). In contrast, poorer concordance was observed for other antibiotics (folate pathway antagonists, polymyxins, tetracyclines, and aminoglycosides; kappa <0.4).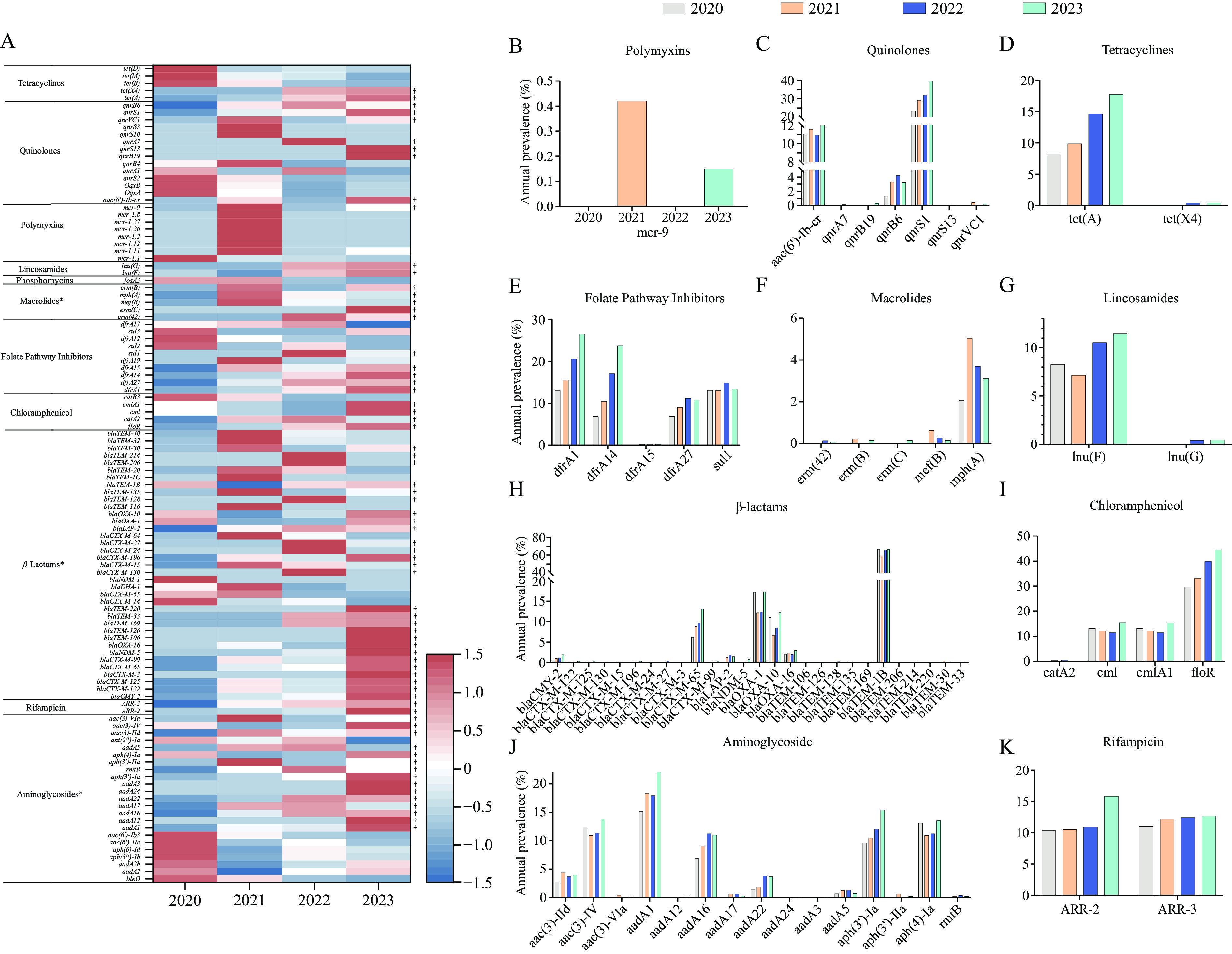 Figure 1.
Figure 1.Temporal distribution of resistance genes detected in Salmonella 4,[5],12:i:- during 2020-2023. (A) The proportion of each gene detected in each year; (B–K) The dynamic prevalence of key antimicrobial resistance genes identified over time. Note: A total of 112 resistance genes (aac(6')-Iaa excluded), encompassing 11 antimicrobial classes, were detected in 2,736 Salmonella 4,[5],12:i:- isolates from 2020-2023. The proportion of each gene was standardized, subjected to Cochran-Armitage trend tests, and visualized with different colors (A), red indicates an increase, and blue indicates a decrease. Three antimicrobial classes (aminoglycosides, beta-lactams, and macrolides) showed significant year-by-year increases. The dynamic change of antimicrobial resistance genes in different classes is shown in B-K. B is polymyxins class, C quinolones, D tetracyclines, E folate pathway inhibitors, F macrolides, G lincosamides, H β-lactams, I chloramphenicol, J aminoglycoside, and K rifampicin.
* indicates a significant increase (Z<0 and P<0.05) in resistance to specific antimicrobial classes;
† indicates a significant increase in the abundance of specific genes over time.
-
To investigate the genetic relationships between isolates, a phylogenetic tree was constructed based on core genomic SNPs for 2,736 Salmonella 4,[5],12:i:- isolates. The analysis revealed five major clades, A–E, with D and E being dominant (Figure 2). Isolates from different years and regions were interspersed throughout these clades, indicating high diversity and genetic heterogeneity of Salmonella 4,[5],12:i:- nationwide. Significant differences in the prevalence of most genes (11/19) were observed between clades (Table 1). Clades A and C contained fewer β-lactamase resistance genes, while most isolates harbored the blaTEM-1B gene. Clades B and E carried more β-lactamase resistance genes, including various subtypes of blaTEM. Clade D contained the greatest number of blaCTX-M-65 and blaOXA genes (blaOXA-1, blaOXA-10, blaOXA-16), substantially more than other clades (Figure 2, Table 1), identifying clade D as a “super-resistant” clone. The strains in clade D were widely distributed across 28 provinces between 2017 and 2023, with the highest number originating from Zhejiang Province (163/897). No clusters (more than 3 isolates with ≤5 SNPs) were detected in the phylogenetic tree.
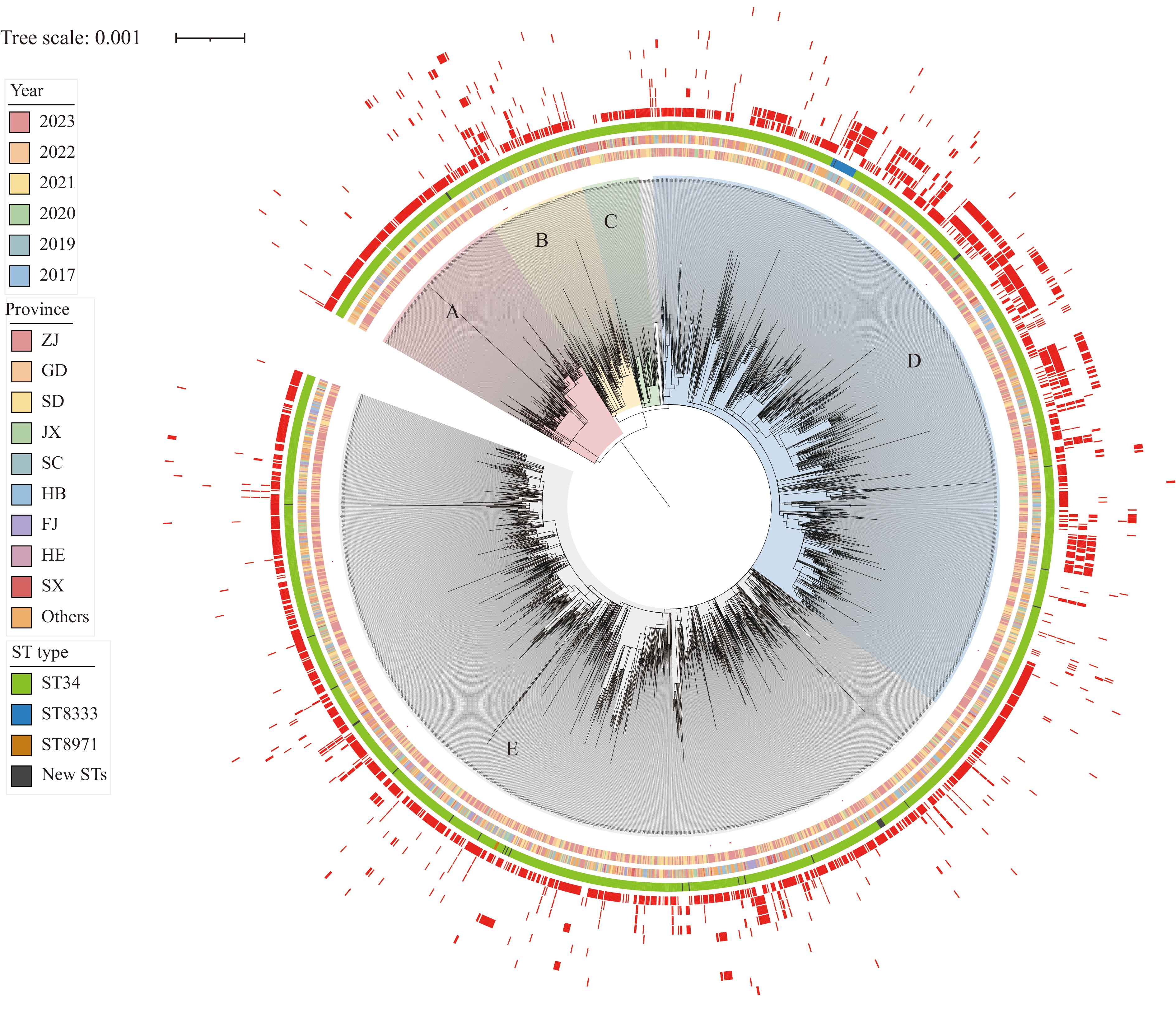 Figure 2.
Figure 2.Phylogenetic tree of Salmonella 4,[5],12:i:- isolates in China from 2017 to 2023 based on core-genome SNPs.
Note: A maximum likelihood tree of Salmonella 4,[5],12:i:- was constructed based on core-genome SNPs. To clearly elucidate relationships, strains on long branches were removed, resulting in a tree containing 2,413 strains. It showed that all strains formed five major clades, A–E, with each clade exhibiting different characteristics, particularly in antimicrobial resistance genes. From inside to outside: year of collection, location, ST type, and resistance genes (in order: blaTEM-1B, blaOXA-1, blaCTX-M-65, blaOXA-10, blaCTX-M-55, blaOXA-16, blaCTX-M-14, blaCMY-2, blaLAP-2, blaDHA-1, blaNDM-5, other β-lactam resistance genes with fewer than 10 isolates each). Carbapenem-resistant isolates are labeled with red dots near the strain code.
Abbreviation: SNP=single-nucleotide polymorphism.
Resistance gene Clade A (n=193) Clade B (n=118) Clade C (n=69) Clade D (n=897) Clade E (n=1,136) P blaCTX-M genes* 4.66 21.19 1.45 28.43 11.88 <0.001 blaCTX-M-14 0.52 9.32 0 1.34 2.38 <0.001 blaCTX-M-55 1.55 4.24 0 3.57 3.87 0.247 blaCTX-M-65 1.55 12.71 1.45 23.30 5.37 <0.001 blaCMY-2 3.11 2.54 0 2.68 0.53 <0.001 blaOXA genes† 1.55 12.71 1.45 23.30 5.46 <0.001 blaOXA-1 2.07 16.95 2.90 31.22 7.04 <0.001 blaOXA-10 0.52 15.25 1.45 19.51 5.46 <0.001 blaOXA-16 0 0 0 7.80 0 <0.001 blaNDM-5 1.04 5.08 0 0.11 0.26 <0.001 blaDHA-1 0 0 1.45 0.33 0.53 0.455 blaLAP-2 0.52 2.54 0 1.45 1.85 0.497 blaTEM genes§ 91.19 66.95 50.72 49.94 75.44 0.497 blaTEM-1B 89.64 66.95 50.72 49.61 74.82 <0.001 blaTEM-1, sul2 and tet(B)¶ 10.88 50.00 43.48 32.78 60.05 <0.001 aac(6’)-Ib-cr 2.07 2.54 2.90 5.24 22.45 <0.001 qnrS1 88.08 30.51 2.90 45.93 11.80 <0.001 qnrS2 0 0 0 3.34 1.23 <0.001 qnrB4 0 0 1.45 0.33 0.53 0.455 qnrB6 0.52 0 0 0.56 7.83 <0.001 qnrB19 0.52 0 0 0.11 0.18 0.564 qnrA1 0.52 0 0 0 0.09 0.290 qnrVC1 0 0 0 0.56 0 0.109 mcr-1 0 0 0 0.11 0.53 0.489 mcr-9 0 0 0 0.22 0.09 0.753 Note: Statistical significance was determined using p-values, where P≤0.05 was considered statistically significant.
* Including all blaCTX-M subtypes present in Salmonella 4,[5],12:i:- genomes in this study.
† Including all blaOXA subtypes present in Salmonella 4,[5],12:i:- genomes in this study.
§ Including all blaTEM subtypes present in Salmonella 4,[5],12:i:- genomes in this study.
¶ Simultaneous presence of blaTEM-1, sul2, and tet(B) genes.Table 1. Association between phylogenetic clades and the presence of resistance genes in Salmonella 4,[5],12:i:- isolates collected in China, 2017–2023.
In addition, three major sequence types were identified among the Salmonella 4,[5],12:i:- isolates. The vast majority (97.33%, 2,664/2,736) belonged to ST34, while 26 isolates were ST8333 (predominantly from Sichuan Province, 13/26), and 3 were ST8971, with an additional 43 novel STs. Although 19 of the 26 ST8333 strains simultaneously carried blaOXA-1, blaCTX-M-65, and blaOXA-10 genes, the limited number of these strains precludes a definitive assessment of whether they carry more resistance genes than ST34 or other sequence types. Among the 2,664 Salmonella 4,[5],12:i:-ST34 sequences, none possessed genetic determinants contributing to the ASSuT profile, whereas 40.77% (1,086/2,664) exhibited the ASuT profile and 16.70% (445/2,664) harbored genetic determinants (blaCTX-M and blaCMY) conferring resistance to extended-spectrum cephalosporins.
-
Fifteen carbapenem-resistant isolates were found to co-harbor the blaNDM-5 gene along with 5-19 additional resistance genes, including blaOXA-10, blaOXA-1, qnrS1, sul3, cmlA3, and floR. Among the eight completed genomes, six contained blaNDM-5 and blaOXA-10 co-existing on IncHI2/IncHI2A plasmids (149,758bp-263,737bp), one carried blaNDM-5 and blaOXA-1 on a p0111 plasmid (106,847 bp), and the remaining isolate harbored blaNDM-5 and blaOXA-1 on an IncI1-I (Alpha) plasmid (123,291 bp). The genes blaNDM-5 and blaOXA-10 were associated with distinct insertion elements (IS3000 and IS1), indicating that these two resistance genes were acquired through independent recombination events. Additionally, these plasmids carried multiple other antimicrobial resistance genes conferring concurrent resistance to carbapenems, third-generation cephalosporins, ceftazidime/avibactam, chloramphenicol, tetracycline, and streptomycin. Comparison of a representative plasmid, pSa23800-2 (Figure 3A), with the NCBI database revealed that the plasmid with highest similarity was pNDM33-1, isolated from an E. coli strain from ducks in Guangzhou, China, in 2020 (Figure 3B). This finding suggests that resistance plasmids may be horizontally transferred between different bacterial species with food serving as a potential vehicle. The second highest similarity was observed with plasmid pCFSAN086837 from S. Newport isolated from poultry in Vietnam in 2017 (Figure 3B), suggesting that these plasmids can persist across different Salmonella serotypes and that human-derived plasmids may originate from animal sources. Furthermore, two carbapenem-resistant isolates (Sa23810 and Sa23811) from Guangdong Province recovered in 2023 exhibited close genetic relationships, with only 6 SNPs differentiating them, while no SNP differences were observed between two other isolates (Sa23800 and Sa23801) from the same year. These findings indicate that two distinct clusters or potential outbreaks occurred in Guangdong Province in 2023.
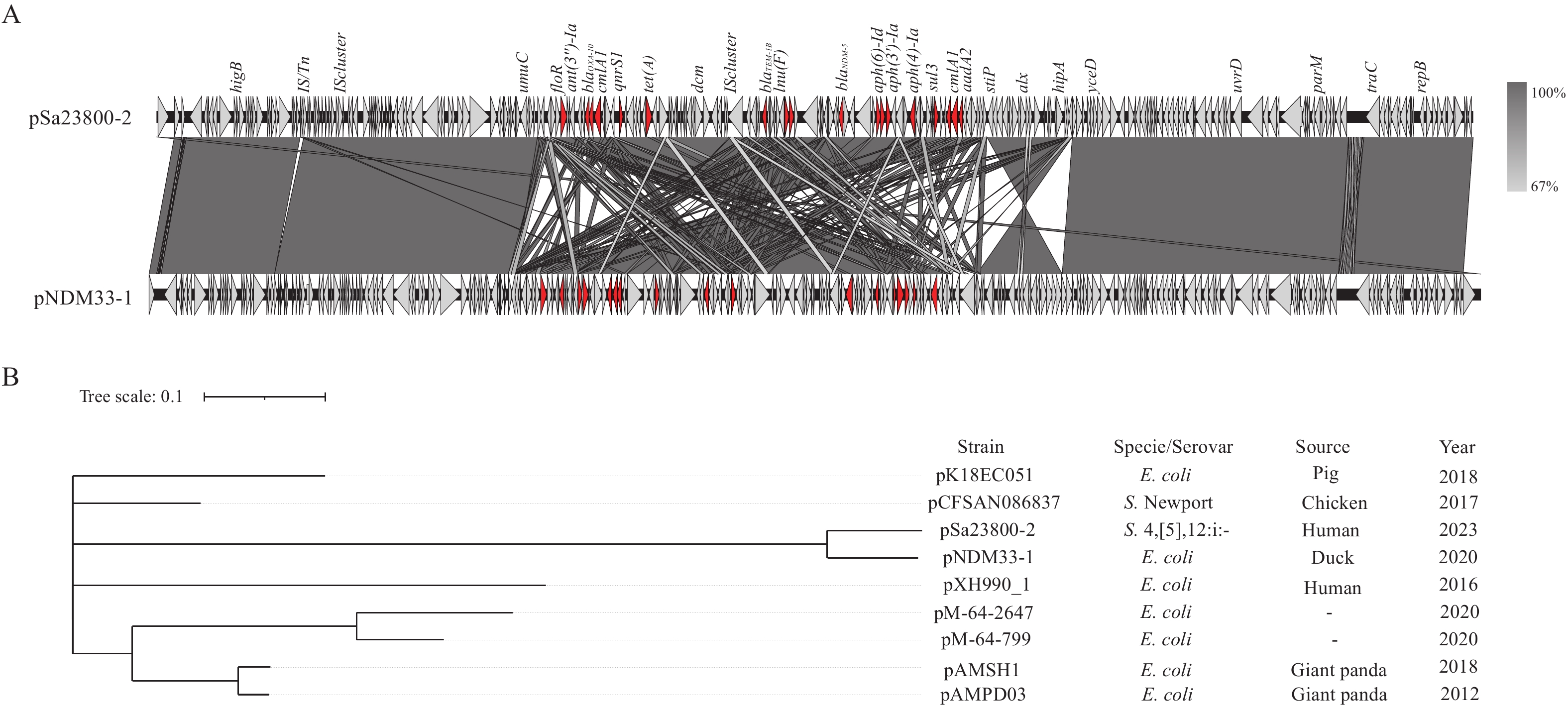 Figure 3.
Figure 3.Alignment and dendrogram of the representative plasmid pSa23800-2 mediating carbapenem resistance. (A) Genomic structure and alignment of plasmid pSa23800-2 with pNDM33-1; (B) Dendrogram of pSa23800-2 and other similar plasmids from various sources.
Note: The gene blaNDM-5 is carried by multiple plasmids conferring carbapenem resistance in Salmonella 4,[5],12:i:-. As a representative, plasmid pSa23800-2, which was used as a query for an NCBI BLAST search, was compared with the highest similarity plasmid, pNDM33-1, from E. coli (3A), and the reference tree was constructed by combining similar plasmids from various sources (3B). The plasmid backbone and virulence genes are colored gray, and AMR determinants are colored red. The areas between ORFs denote nucleotide sequence identity. -
This study provides a comprehensive nationwide assessment of antibiotic resistance in human-derived Salmonella 4,[5],12:i:-, a predominant serovar causing human Salmonella infections in China as determined through systematic surveillance (7). We elucidated its evolutionary history using a large genomic dataset collected over seven years across numerous provinces. Among these isolates, 66% were from children under 5 years of age, confirming that this specific serovar predominantly affects infants, consistent with previous reports (20-21). Our analysis revealed exceptionally high resistance rates to streptomycin, tetracycline, and ampicillin (all >85%) in human-derived Salmonella 4,[5],12:i:-, aligning with previous findings (13,22-23). Notably, resistance to third-generation cephalosporins is significantly higher in China than in developed countries in Europe and America (22% vs. 5%). For the first time, we identified numerous Salmonella 4,[5],12:i:- isolates resistant to carbapenems and the "last-resort" antibiotic colistin, potentially indicating inappropriate use of these antibiotics in clinical treatment and animal food production. Widespread carbapenem resistance could lead to increased treatment failures, higher mortality rates, greater healthcare resource burdens, longer hospital stays, and increased costs. This situation may compromise the effectiveness of empiric treatment regimens, necessitate more invasive diagnostic procedures, delay appropriate therapy, and require more expensive alternative treatments, disproportionately affecting vulnerable populations with limited healthcare access. Over the four-year study period, resistance to key antimicrobials (meropenem, azithromycin, and ampicillin/sulbactam) continued to increase, while resistance to classic first-line antibiotics (ampicillin, streptomycin, and tetracycline) remained consistently high. The predominant AST resistance profile, which differs from the ASSuT profile commonly observed in Europe and America (11-12), suggests widespread use of tetracycline, penicillins, and aminoglycosides in food animals in China in recent decades, with resistance potentially linked to extensive use of these drugs in food-producing animals. Importantly, the increased antibiotic resistance levels of Salmonella 4,[5],12:i:- may contribute to the further spread of this serovar, highlighting the importance of establishing targeted nationwide surveillance using a One Health approach to determine the prevalence and spread across different sources and implement appropriate control or risk mitigation measures.
The ST34 genotype of Salmonella 4,[5],12:i:- has been the dominant epidemic type over the past few decades (11,12,24); similarly, in our study, ST34 was the most common genotype, along with ST8333 and ST8971. The epidemic success of Salmonella 4,[5],12:i:- ST34 is partially attributed to the selective acquisition of antibiotic resistance (4,25–26), leading to the establishment of MDR clones (4). Our analysis detected numerous antibiotic-resistance genes located in both chromosomes and mobile genetic elements that have accumulated in Salmonella 4,[5],12:i:- (Figures 1 and 2), particularly in clade D of ST34, where ESBL genes (i.e., blaCTX-M, blaOXA-10) were enriched. Additionally, we identified multiple plasmid-carried carbapenemase genes (i.e., blaNDM-5 and blaNDM-1) conferring carbapenem resistance in 15 ST34 Salmonella 4[5],12:i:- isolates. Carbapenem-resistant Salmonella was first identified in humans in the United States in 2011 (in a patient with a history of medical treatment in India) (27). In China, blaNDM-1-carrying Salmonella was initially detected in 2012 in a stool sample from a child with diarrhea in Zhejiang Province (28). Since then, carbapenem-resistant Salmonella (such as S. Typhimurium, S. Indiana, S. London, and S. Stanly) has been reported in sporadic cases or carriers (29–32). The resistance genes involved were mainly blaNDM-5 and blaNDM-1 (with occasional presence of blaKPC-2 or blaOXA-48 or coexistence of mcr-1, blaCTX-M-65, and blaOXA-1), predominantly carried by transferable plasmids (such as IncX3, IncHI2, and IncFIB). However, no Salmonella 4,[5],12:i:- isolate coharboring blaNDM-5 and blaOXA-10 has been previously reported. Our study is the first to identify a large number of strains carrying IncHI2/IncHI2A plasmids that co-harbor blaNDM-5, blaOXA-10, and other AMR genes, indicating the emergence of novel multidrug-resistant subclones (some extensively drug-resistant) in Salmonella 4,[5],12:i:- ST34. The IncHI2 plasmid is a type of conjugative plasmid widely found in Salmonella enterica that mediates MDR (20,33–34). The presence of blaNDM and blaOXA genes carried by IncHI2-like plasmids highlights the widespread horizontal exchange of plasmids between Salmonella 4,[5],12:i:- and different bacterial species, emphasizing the necessity of monitoring and understanding the transmission and genetic characteristics of bacterium-plasmid pairs. Additionally, the abundance of resistance genes, profiles, and variant plasmids carried by Salmonella 4,[5],12:i:- highlights the genomic plasticity of this serovar. Despite the low rate of resistance to both carbapenems and colistin (≤1%) in Salmonella 4,[5],12:i:-, the sharp increase in the number of isolates resistant to these two agents in 2023, along with resistance rates to third-generation cephalosporins exceeding 20%, is highly concerning because high levels of third-generation cephalosporin resistance may drive increased use of carbapenems and/or the ‘last-resort’ antibiotic colistin.
The widespread use of antibiotics in food-producing animals, especially those less commonly used in humans, such as enrofloxacin and florfenicol, may lead to the emergence of resistant strains. These strains can enter humans through the food chain or environmental contamination, contributing to the development and accumulation of antibiotic resistance (35). This was corroborated by our detection of certain resistance genes in Salmonella 4,[5],12:i:-, such as floR (florfenicol resistance), qnrS1, qnrB6, and qnrB14 (quinolone resistance), and their increasing prevalence over the study period. The global epidemic ST34 clone (ASSuT profile) circulating in Europe was introduced into the swine sector in the U.S. Midwest from the EU in 2014 (11−12) and is widespread, being detected in humans, farms, and the environment, but remains predominantly associated with the pig reservoir. However, the absence of the ASSuT profile of ST34 in our study may indicate distinct endemic clone expansion and circulation in China. To some extent, the high similarity of drug resistance plasmids in Salmonella 4,[5],12:i:- with those in other Salmonella serotypes and species (i.e., pSa23800-2 in humans and pNDM33-1 in ducks) partially reveals the occurrence of regional cross-contamination and suggests zoonotic transmission. This zoonotic transmission highlights the importance of considering animal populations in the overall strategy to control the spread of antibiotic-resistant bacteria. The movement of resistant strains between animals and humans can facilitate the dissemination of resistance genes across different niches or ecosystems. Further investigations, including a time-scaled phylogeny with a discrete trait geospatial model and epidemiological information associated with the cases, are needed to determine the origin and transmission routes of Salmonella 4,[5],12:i:- in China.
Moreover, the phylogenetic tree revealed that Salmonella 4,[5],12:i:- strains with diverse resistance gene profiles were interspersed throughout China, with no distinct clusters identified. However, highly similar strains were observed among carbapenem-resistant isolates, suggesting both high genetic diversity and potential clonal transmission of carbapenem resistance, which warrants vigilant monitoring. Different antimicrobial resistance genes were acquired by distinct clades/lineages of Salmonella 4,[5],12:i:-, enabling the emergence of multiple clones with resistance to various antibiotic families — clade D with ESBL genes, clade B with blaNDM-5, clade E with aac(6')-Ib-cr, and clade A with qnrS1 (Table 1). Two variants of mcr genes were identified in 13 colistin-resistant Salmonella 4,[5],12:i:- ST34 strains. The mcr-1 gene, first reported in China (36), has predominantly spread across human, pig, and chicken populations via conjugative plasmids in Asia, Europe, and Africa, particularly in China, Portugal, and England (26), suggesting that Asia may serve as a potential reservoir for global dissemination. Similarly, mcr-9.1, the second most common variant of the mcr gene (after mcr-1.1) identified in Salmonella 4,[5],12:i:-, is also globally widespread across all sectors (37) and can be horizontally transferred via the IncHI2 plasmid (38). The more frequent detection of blaNDM, mcr, and ESBL genes in Salmonella 4,[5],12:i:- compared to other serotypes of Salmonella enterica indicates that this serovar may more readily acquire exogenous genes and serve as a potential reservoir of resistance genes.
This study had several limitations. The strains originated primarily from eastern China, with a limited number from other regions, which may introduce some bias into the results. However, there was no significant difference in the Salmonella spp. isolation rate among different regions in China. Secondly, the strains lacked clinical metadata, such as detailed patient outcomes and exposure history, which could provide deeper insights into the factors influencing resistance patterns and transmission. In addition, the study focuses only on human-derived isolates and does not include environmental or animal sources.
In conclusion, this retrospective analysis of the antimicrobial resistance phenotypes and genetic characteristics of human-derived Salmonella 4,[5],12:i:- isolates collected between 2017 and 2023 reveals widespread and significant resistance in China. High-level resistance to third-generation cephalosporins and the emergence of over 0.5% carbapenem resistance, along with the presence of plasmid-carried genes for resistance to key antimicrobial classes (such as blaNDM and ESBL genes in certain lineages of Salmonella 4,[5],12:i:-), which may become a new epidemic MDR/XDR clone in China, highlight the need for enhanced surveillance of resistance genes, clones, and serovar-plasmid pairs. ESBL-producing, carbapenem-resistant, and colistin-resistant strains are usually multidrug-resistant and are often associated with treatment failure. Therefore, further investigations to determine the factors explaining the overall emergence of resistance in recent years are crucial for preventing the spread of Salmonella strains bearing resistance genes and novel resistant clones, particularly in targeted young populations.
HTML
Data Source and Bacterial Strains
Antimicrobial Susceptibility Testing
Phylogenetic Analysis
Statistical Analysis
Increasing Resistance Trend and Emergence of Carbapenem Resistance in Salmonella 4,[5],12:i:-
Salmonella 4,[5],12:i:- Exhibited a Substantial Number of Resistance Genes, the Prevalence of Which Increased Annually
High Genetic Heterogeneity and Diversity of Salmonella 4,[5],12:i:- in China, with Clade D Carrying More Cephalosporin Resistance Genes
Most Multidrug Resistance Plasmids Harbor blaNDM-5 in Carbapenem-Resistant Salmonella 4,[5],12:i:- and are Distributed across Different Bacterial Species
| Citation: |

|