
 Download:
Download:




-
Streptococcus agalactiae, classified as a group B streptococcus (GBS) (1), can exist as a commensal organism in the human gastrointestinal and genitourinary tracts while also causing severe infections in immunocompromised individuals and the elderly. In pregnant women, GBS infection is associated with maternal disease (2) and can cause early-onset or late-onset sepsis in newborns and hypoxic-ischemic encephalopathy in infants (3). The global prevalence of maternal GBS colonization is estimated at 18%, with significant regional variations. The pooled incidence of invasive GBS disease in infants is 0.49 per 1,000 live births, highest in Africa (1.12) and lowest in Asia (0.30) (4–5). In China, the maternal colonization rate is 8.1%, lower than the global average (6). GBS also represents a growing threat to immunocompromised individuals and the elderly (7).
Intrapartum antibiotic prophylaxis (IAP) effectively prevents GBS infections, but overuse of antibiotics contributes to increasing antibiotic resistance in GBS (8). Additionally, antibiotic exposure in newborns can have detrimental health effects (9). These concerns highlight the need for alternative treatment approaches, such as GBS vaccines. Capsular polysaccharides (CPs) are key virulence factors in GBS, with 10 recognized serotypes (Ia, Ib, II-IX) based on their immunological reactivity (10). Understanding the distribution of these serotypes is essential for the development of effective CPS-based vaccines (4). Currently, no vaccine is available for GBS infection. Clarifying GBS serotype distribution remains critical for developing effective prevention and treatment strategies, but studies in China are limited in scope and geographic coverage (6,11–12).
This study provides an extensive genomic characterization of GBS strains collected in China, aiming to investigate serotype and genotype distribution, genetic evolution, and the presence of virulence and antibiotic resistance genes. By elucidating the molecular and genomic epidemiological patterns of GBS in China, this research establishes a foundation for developing effective prevention and treatment strategies.
-
A total of 747 Streptococcus agalactiae genomes were analyzed in this study, comprising 721 publicly available genomes retrieved from the National Center for Biotechnology Information (NCBI) database (https://www.ncbi.nlm.nih.gov/datasets/genome/) as of April 2024, supplemented with 26 newly sequenced clinical isolates. The clinical strains were sequenced using the Illumina HiSeq 2000 platform (Novogene Technology Co., Ltd., Beijing, China), and genome annotation was performed using Prokka [version 1.13.3; Winnipeg Institute for Bacterial Genomics (PHAC), Winnipeg, Canada]. The basic information of all strains is provided in
Supplementary Table S1 . -
Serotype determination was conducted through CPs sequence analysis using pan-genomic comparison, with a threshold of ≥95% sequence identity required for serotype confirmation (13–14). Phylogenetic analyses were performed based on core single nucleotide polymorphisms (SNPs) using IQ-TREE (version 2.3.4; Computational Evolution Lab, Ho Chi Minh City, Vietnam). Virulence and antibiotic resistance genes were identified from Prokka-generated *.gff files using Roary (version 2.3.12; University of Exeter, Exeter, UK). Virulence genes were identified through BLASTp alignment against the Virulence Factors of Pathogenic Bacteria (VFDB, https://www.mgc.ac.cn/VFs/) database, applying stringent criteria (≥60% identity, ≥70% coverage, E-value ≤1×10-5). Antibiotic resistance genes were identified using the Resistance Gene Identifier (RGI) [version 6.0.2; Comprehensive Antibiotic Resistance Database (McMaster University), Hamilton, Canada] against the Comprehensive Antibiotic Resistance Database (CARD, https://card.mcmaster.ca/home), with thresholds of ≥80% for both identity and coverage.
-
Statistical analyses were conducted using SPSS (version 26.0; IBM Corp., Armonk, NY). Categorical variables were assessed using Fisher’s exact test, with statistical significance defined as P<0.01. This test was specifically employed to evaluate differences in serotype distribution of GBS across various geographical regions of China. The genome sequences of the 26 newly sequenced clinical GBS strains have been deposited in GenBank under BioProject ID PRJNA1128393.
-
A total of 747 Streptococcus agalactiae (GBS) strains collected in China from 1998–2024 were classified into eight distinct serotypes (Table 1). Five predominant serotypes (III, Ib, Ia, V, and II) accounted for 96.65% of all isolates, with serotype III being the most prevalent (n=323, 43.24%), followed by Ib (n=177, 23.69%) and Ia (n=120, 16.06%). Serotypes IV (n=4, 0.54%), VII (n=1, 0.13%), and non-typeable strains (n=4, 0.54%) were comparatively rare. The isolation source remained undocumented for over half of the samples (n=428, 57.30%). Among strains with documented sources, blood specimens were the most common origin for serotype III isolates, while genital secretions predominated in serotypes Ib and V.
Serotype Isolation source Total (%) Blood Genital secretion Vaginal and rectal Urine Sputum Others Unknow III 59 8 20 8 4 34 190 323 (43.24) Ib 9 34 6 6 7 13 102 177 (23.69) Ia 11 4 2 2 6 28 67 120 (16.06) V 3 15 2 5 3 12 41 81 (10.84) II 2 1 0 2 1 3 12 21 (2.81) VI 1 1 1 1 0 1 11 16 (2.14) IV 0 0 0 1 0 2 1 4 (0.54) VII 0 0 0 0 0 0 1 1 (0.13) NT 0 0 0 0 0 1 3 4 (0.54) Total 85 63 31 25 21 94 428 747 (100) Note: Columns 1 to 5 under isolation source represent the primary sample types, all of which were collected from humans. Others: Include cervical samples, milk, skin, fish farm, bovine sources, and additional specimen types.
Abbreviation: GBS=group B Streptococcus.Table 1. Serotype and isolation source distribution of GBS strains in China.
-
Spatio-temporal analysis revealed a low prevalence of GBS from 1998 to 2011, followed by a significant increase starting from 2012 (Figure 1A). Serotype III has consistently been the predominant strain, while serotype Ib has shown a steady increase since 2016. Serotypes V and Ia have maintained relatively stable frequencies at lower levels (Figure 1B). Among the 623 strains with available regional data, the majority originated from Taiwan, China (35.3%); Shandong Province (18.6%); Hong Kong Special Administrative Region (SAR, 8.4%); and Henan Province (8.4%). Serotype III demonstrated clear dominance in Taiwan, China (58.71%) and Hong Kong SAR (60.32%), whereas serotype Ib was most prevalent in Shandong Province (48.20%), Henan Province (53.97%), and Shanghai Municipality (44.44%). Statistical analysis revealed significant regional variations in serotype distribution (P<0.01, Table 2).
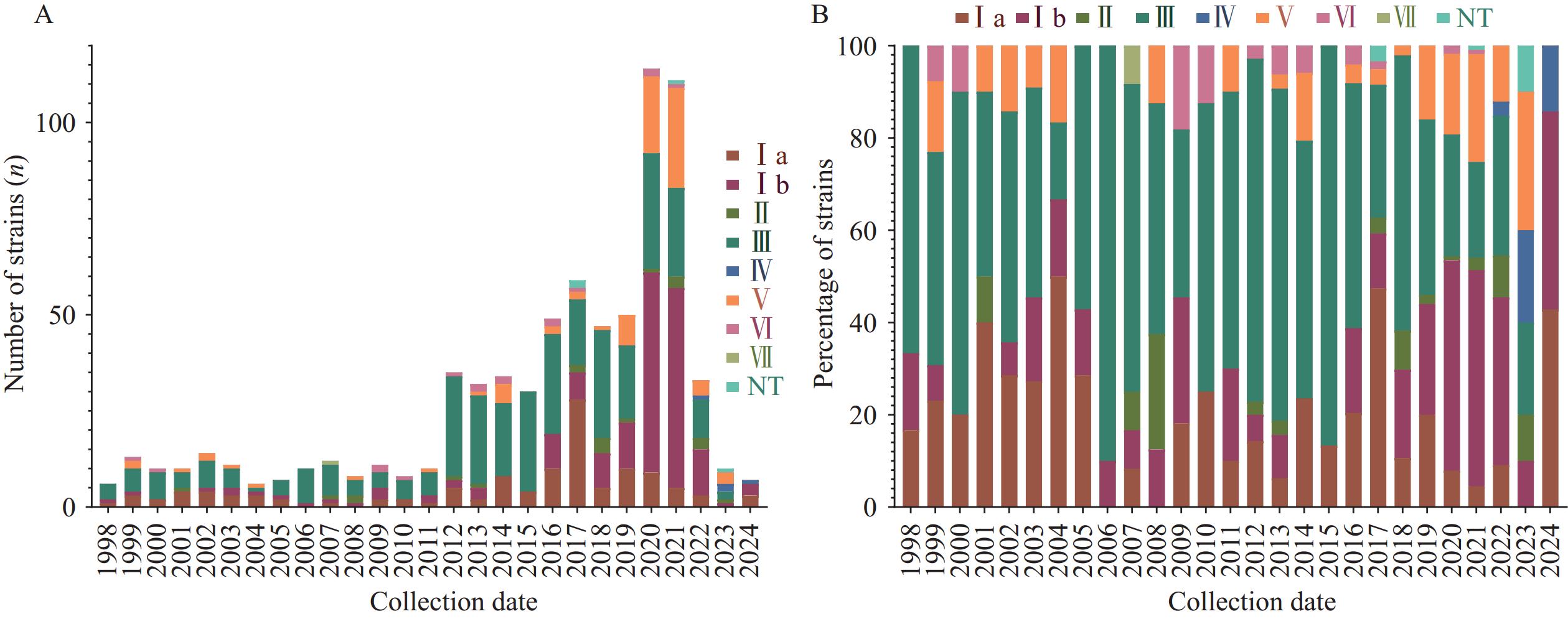 Figure 1.
Figure 1.Spatio-temporal distribution of GBS in China. (A) Temporal distribution of GBS in China; (B) Temporal distribution of GBS in China, percentage stacked bar chart.
Abbreviation: GBS=group B Streptococcus.Region Serotype* Total Ⅰa Ⅰb Ⅱ Ⅲ Ⅳ Ⅴ Ⅵ Ⅶ Ⅷ Ⅸ NT Taiwan, China 51 22 8 155 0 16 11 1 0 0 0 264 Shandong 7 67 2 34 1 25 2 0 0 0 1 139 Henan 4 34 1 8 0 15 1 0 0 0 0 63 Hong Kong SAR 19 1 0 38 0 2 1 0 0 0 2 63 Shanghai 5 20 2 13 0 5 0 0 0 0 0 45 Beijing 2 7 0 6 0 1 0 0 0 0 0 16 Anhui 0 3 3 2 3 3 0 0 0 0 1 15 Guangdong 3 1 0 2 0 0 0 0 0 0 0 6 Jiangsu 1 1 1 0 0 1 1 0 0 0 0 5 Zhejiang 0 0 0 2 0 1 0 0 0 0 0 3 Guangxi 1 0 0 0 0 0 0 0 0 0 0 1 Hainan 1 0 0 0 0 0 0 0 0 0 0 1 Ningxia 1 0 0 0 0 0 0 0 0 0 0 1 Sichuan 0 0 0 1 0 0 0 0 0 0 0 1 Not collected 25 21 5 61 0 12 0 0 0 0 0 124 Total 120 177 22 322 4 81 16 1 0 0 4 747 Abbreviation: GBS=group B Streptococcus; Hong Kong SAR=Hong Kong Special Administrative Region.
* P<0.001.Table 2. Serotype and spatial distribution of GBS strains in China.
-
A maximum likelihood phylogenetic analysis of the 747 GBS genomic sequences revealed five distinct evolutionary lineages (Figure 2A). Lineage 5 comprised the largest group (n=281), predominantly containing serotypes III (n=184) and Ib (n=76). Lineage 3 encompassed 274 strains, with serotype Ib (n=176) being the most prevalent. Lineage 4 contained 141 strains, primarily consisting of serotypes III (n=93) and V (n=44). Lineage 2 was notably smaller, containing only two strains of serotype II, isolated from bovine sources in 2021. The analysis revealed no significant direct correlation between serotype classification and genetic evolutionary patterns, nor was there evidence of a relationship between phylogenetic lineage and endemic circulation patterns of the strains.
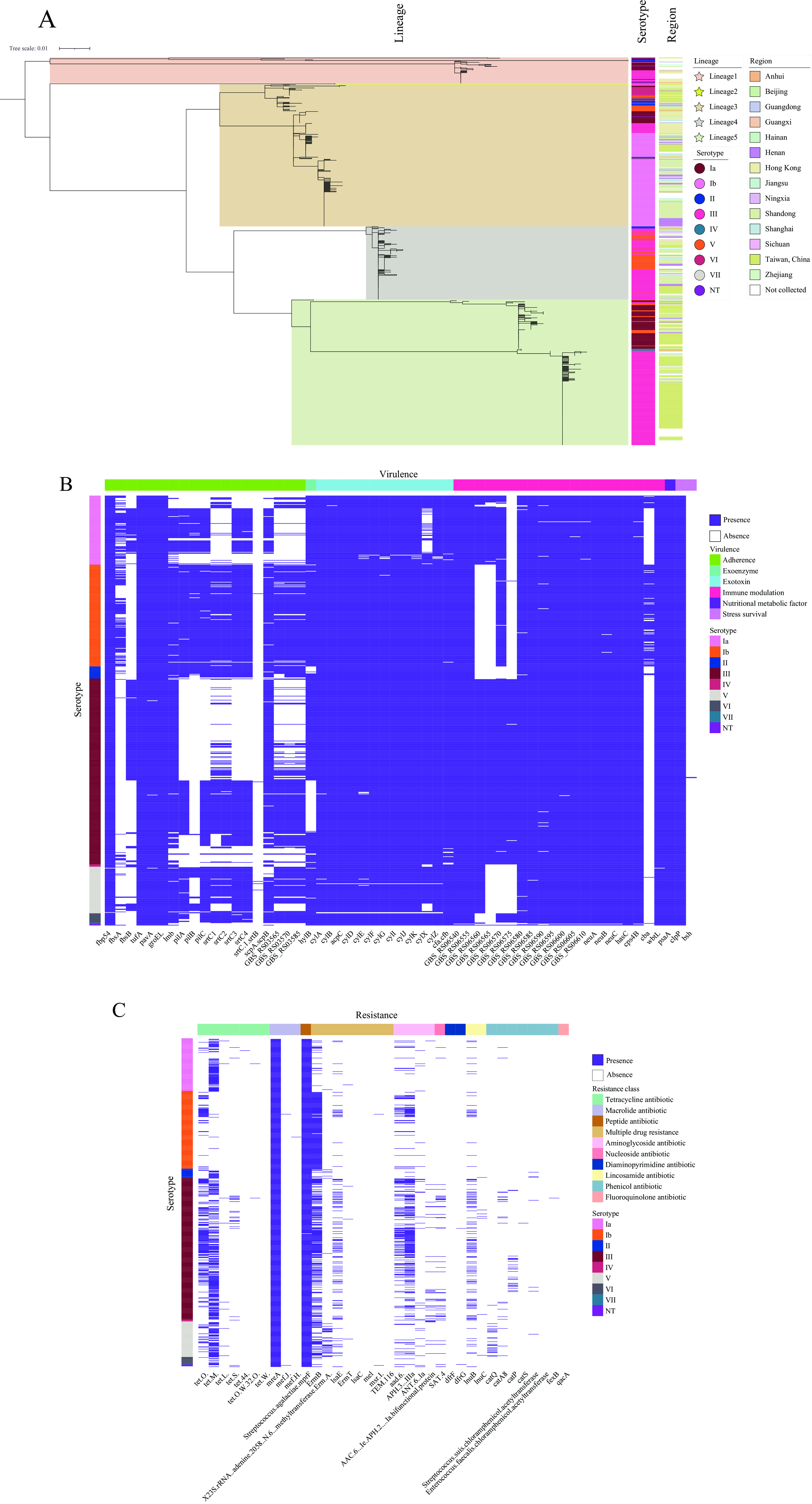 Figure 2.
Figure 2.Phylogenomic tree and distribution of virulence- and ARGs-associated genes across 747 Streptococcus agalactiae genomes. (A) Phylogenomic tree of 747 GBS genomes constructed using maximum likelihood method based on core gene sequences; (B) Distribution of virulence-associated genes across 747 GBS genomes; (C) Distribution of antibiotic resistance genes across 747 GBS genomes.
Note: Genes with comparable functions or those belonging to the same functional class have been integrated in the visualization.
Abbreviation: GBS=group B Streptococcus.
-
A comprehensive analysis of 747 GBS genomes identified 56 virulence-associated genes, categorized into six functional groups: adherence, exoenzyme, exotoxin, immune modulation, nutritional/metabolic factors, and stress survival (Figure 2B, Table 3). Each genome harbored an average of 45 virulence genes, with 33 genes present in nearly all strains. While most strains carried either fbsA or fbsB, 161 strains (predominantly serotypes III and V) lacked these fibrinogen-binding genes. Notably, 48 serotype Ia and 205 serotype III strains lacked genes associated with bacterial pili (pilA, pilB, pilC, setC1, srtC2, srtC3, srtC4, GBS_RS03565, GBS_RS03570, GBS_RS03585), while 92 serotype III strains lacked the hyaluronidase gene (hylB). Additionally, 60 serotype Ia strains lacked the cylX gene, and strains of serotypes Ib (n=177), II (n=21), and V (n=81) were less likely to carry genes related to the GBS capsule. The cba gene was predominantly found in serotypes Ia (n=20) and Ib (n=148).
VF category Virulence genes Virulence factor Functions Adherence tufA, groEL, fbp54, pavA, lmb, scpA/scpB, srtC-1/srtB, srtC3, srtC4, pilC, pilA, GBS_RS03585, srtC2, GBS_RS03565, GBS_RS03570, srtC1, pilB, fbsB, fbsA EF-Tu, GroEL, FBPs, PavA, Lmb, C5a peptidase, PI-2a, PI-1, FbsB, FbsA, RlrA islet Adherence and colonization; promotes fibronectin-mediated collagen recruitment, adhesion and invasion; dissemination; impairs recruitment to sites of infection; biofilm formation; protein binding, recognizing the extracellular matrix; manipulates the host inflammatory response Exoenzyme hylB Hyaluronidase Dissemination, penetration Exotoxin acpC, cylD, cylB, cylF, cylI, cylJ, cylA, cylK, cylZ, cylG, cfa/cfb, cylE, cylX Beta-haemolysin/cytolysin, CAMP factor Forms pores in cell membrane; pro-inflammatory effects: induces apoptosis, promotes cellular invasion, and triggers iNOS and cytokine release Immune modulation hasC, GBS_RS06540, GBS_RS06555, neuA, neuC, wbtL, GBS_RS06585, GBS_RS06595, GBS_RS06600, GBS_RS06605, GBS_RS06610, neuB, cps4B, GBS_RS06580, GBS_RS06590, GBS_RS06570, GBS_RS06560, GBS_RS06565, GBS_RS06575, cba Hyaluronic acid capsule, Capsule, LPS, Beta-C protein Prevents phagocytosis; discourages C3b binding; binds to CD44, preventing the activation of the alternative complement pathway; inhibits complement-mediated opsonophagocytosis; LPS, resistant to complement deposition, masks cell wall-associated complement, preventing opsonophagocytosis Nutritional/Metabolic factor psaA PsaA Transport of Mn2+ and Zn2+ into bacterial cytoplasm, mutants impacts colonization and increase susceptibility to oxidative damage Stress survival clpP, bsh ClpP, BSH Serine protease involved in proteolysis and resisting the acute toxicity of bile and bile salts Abbreviation: GBS=group B Streptococcus. Table 3. The classification of virulence-related factors of Streptococcus agalactiae.
The genomic analysis revealed 27 antibiotic resistance genes (ARGs) associated with nine distinct antimicrobial drug classes (Figure 2C). Most genomes contained genes conferring resistance to peptides (Saga_mprF) and macrolide antibiotics (mreA), indicating fundamental resistance mechanisms in GBS. Over half of the strains carried genes for tetracycline resistance [tet(M), n=387; serotype III, 210; Ia, 85; V, 61] and erythromycin resistance (ermB, n=429; serotype III, 192; Ib, 173; V, 38). Additionally, substantial proportions of strains carried genes encoding APH3 (32.8%, n=245; serotype III, 176), tetO (29.6%, n=221; serotype III, 151), aad6 (20.6%, n=154; serotype III, 102), lnub (15.8%, n=118; serotype III, 88), and lsaE (15.1%, n=113; serotype III, 83). Serotype III GBS strains consistently harbored the highest number of resistance genes. Detailed resistance profiles are provided in
Supplementary Table S2 . -
Among the 747 GBS strains collected in China from 1998 to 2024, we identified eight serotypes and one non-typeable serotype, with serotypes III, Ib, Ia, V, and II constituting 96.65% of all isolates. This distribution closely aligns with both global (98%) and previously reported Chinese (95.9%) distributions (4,6). Notably, the prevalence of serotype III (43.24%) in our study exceeds the global average (25%) but approximates the previously reported Chinese average (46.4%), reflecting significant regional heterogeneity within China (4,6). These findings suggest that while the overall GBS serotype distribution in China mirrors global trends, substantial intra-national variations exist. Geographic distribution analysis revealed marked regional differences, with Beijing Municipality demonstrating a high proportion of serotype III (38.3%), while Taiwan, China reported higher prevalence of serotype VI (11–12). These observations underscore the pronounced geographical variations in GBS serotype distribution across China, emphasizing the necessity for region-specific surveillance studies. Temporal trend analysis indicates an increase in GBS cases, likely attributable to enhanced clinical awareness and advances in sequencing technologies, although the recent decline in case numbers may reflect the impact of coronavirus disease 2019 (COVID-19) pandemic disruptions or delays in sequencing data submission. Serotype III has maintained predominance throughout the study period, while serotype Ib has shown a notable increase in recent years, highlighting the critical importance of comprehensive serotype distribution data for targeted vaccine development strategies.
We identified six categories of virulence genes through VFDB analysis, comprising a total of 56 genes. The adhesion protein FbsA facilitates GBS binding to fibrinogen, thereby protecting the pathogen from phagocytosis (15), while fbsB enhances GBS invasion into host cells and promotes vaginal colonization (16). Several genes (SrtC1, SrtC2, GBS_RS03565, GBS_RS03570, and GBS_RS03585) are associated with the pili type PI-1 structure. The PI-2a pilus comprises three subunit proteins (PilA, PilB, PilC), whose assembly requires sortases SrtC3 and SrtC4, collectively facilitating GBS invasion and colonization (17-18). While pili structures represent potential vaccine targets, strains lacking pili islands must be considered in vaccine development strategies (19). The gene encoding hyaluronidase (hylB) demonstrated a carriage rate of 85.68%, comparable to the 97.8% reported in Zimbabwean isolates (20). This enzyme enables GBS to breach the placental barrier, ascend from the vagina, and infect the fetus (21), while its absence significantly reduces GBS’s capacity to invade fetal tissue and induce preterm delivery (22).
Exotoxin genes from the cyl gene family, cfa, and cfbs, together with the CAMP factor, are associated with β-hemolysis in GBS, causing tissue damage through lysis of human cells (21). The hemolytic properties of GBS play a crucial role in its pathogenesis and immune evasion mechanisms (23). The universal presence of cytochrome-encoding genes across all GBS strains suggests that targeting cytochrome functionality could represent a promising preventive strategy. Immunomodulatory genes, particularly those related to CPs, are critical virulence determinants, with CP-based vaccines currently under development. Our genomic analysis identified several virulence genes associated with CPs, with GBS_RS06560, GBS_RS06565, GBS_RS06570, and GBS_RS06575 being notably underrepresented in serotypes Ib, II, V, and VI. Comparative analysis of capsular polysaccharide composition between these and other strains may elucidate the functional impact of these genes on the capsular structure. CPs enhance GBS virulence by facilitating immune evasion, inhibiting opsonophagocytosis, and promoting biofilm formation (24). The cba gene, predominantly found in serotypes Ia and Ib, may confer protection against opsonophagocytosis (25). While the virulence genes identified in this study provide valuable insights into GBS pathogenicity in China, they represent only a partial characterization, necessitating additional strain data and experimental validation for comprehensive assessment.
Antimicrobial resistance genes conferring resistance to clindamycin, erythromycin, and tetracycline were prevalent among the analyzed strains, with Saga_mprF specifically associated with daptomycin resistance (26–27). The mreA gene and members of the erm gene family were widely distributed, contributing to macrolide-lincosamide-streptogramin B resistance, particularly against clindamycin and erythromycin (28–29). Among 278 strains lacking erm genes, 16 harbored both lsaE and lnuB; however, their precise resistance mechanisms remain unclear due to limited sample availability. Six distinct tet genes were identified, with tetM and tetO encoding ribosomal protection proteins that confer tetracycline resistance (30). Additionally, some strains carried the SAT-4 gene, which confers resistance to nucleoside antibiotics. While intrapartum antibiotic prophylaxis primarily targets early-onset GBS disease, the resistance profiles of GBS isolates continue to evolve, with increasing resistance to beta-lactams, erythromycin, and clindamycin, alongside emerging vancomycin-resistant strains (31). Comprehensive characterization of resistance gene distribution provides valuable insights into GBS resistance patterns, thereby informing the development of effective prevention and treatment strategies.
-
This study provides a comprehensive genomic characterization of Streptococcus agalactiae (GBS) in China, encompassing serotype distribution, virulence factors, and antimicrobial resistance determinants. While the serotype distribution in China generally aligns with global patterns, significant regional variations exist that warrant further investigation. This research contributes essential knowledge for developing effective prevention and treatment strategies for GBS infections in China. Future studies should focus on more detailed regional and temporal analyses to inform targeted prevention approaches, treatment protocols, and vaccine development strategies.
HTML
Bacterial Strains, Whole-Genome Data Collection, and Sequencing
Identification of Serotype, Virulence, and Antibiotic Resistance-Associated Genes
Statistical Analysis and Data Availability
Serotype and Isolation Source Distribution
Spatio-Temporal Distribution
Phylogenetic Tree Construction
Distribution Pattern of Virulence- and ARGs- Associated Genes
| Citation: |

|