
 Download:
Download:




-
Over the past two decades, enterococci have emerged as important multidrug-resistant (MDR) pathogens, responsible for an increasing number of nosocomial and community-acquired invasive infections worldwide (1). The injudicious use of antibiotics in food animal production and human disease treatment has contributed to the spread of MDR enterococci in food chains (2). Enterococcus faecalis (E. faecalis) and Enterococcus faecium (E. faecium) are the two most prevalent enterococcal species identified in human infections (3). Other species, such as Enterococcus casseliflavus (E. casseliflavus), Enterococcus durans, Enterococcus gallinarum (E.gallinarum), and Enterococcus hirae (E. hirae), can also infect humans with concurrent hematological malignancies, neutropenia, and prior corticosteroid treatment (4). Identifying enterococci at the species and molecular levels is clinically relevant due to the antibacterial resistance (ABR) profiles of different isolates. E. faecium exhibits higher rates of ampicillin and vancomycin resistance than E. faecalis (5). E. gallinarum and E. casseliflavus intrinsically exhibit low-level vancomycin resistance (6). E. faecalis and E. faecium hospital-adapted sequence type (ST) and clonal complex (CC) strains (e.g., E. faecalis ST6 and CC2, and E. faecium CC17), and community-adapted lineages (e.g., E. faecium CC94 and E. faecalis ST16) have been identified (7).
The One Health approach requires integrated analysis of important bacteria from animal, food, environmental, and human sources to characterize their populations and resistance phenotypes. Given the complexity of enterococcal ABR, highlighting the One Health approach’s role in addressing this challenge is particularly important (8). Therefore, this study aimed to understand the population structure of MDR enterococcal isolates and their potential transmission risks along the food chain in China.
-
This study was performed in accordance with protocols approved by the Ethics Committee of the State Key Laboratory of the China National Centre for Food Safety Risk Assessment (CFSA).
We collected 694 samples from various sites and sources along the food chain (animal, food, environment, and human) representing the enterococcal population during 2015–2022. Briefly, the samples were collected from livestock (n=224), the farm environment (n=145), retail meat (n=91), retail fruits and vegetables (n=100), the urban environment (n=47), and humans (n=87) in Beijing Municipality, and Hubei, Henan, and Jilin Provinces. The samples collected from Henan and Jilin Provinces covered the entire food chain, including the community population and inpatients with diarrhea. The sample locations, types, and numbers are shown in
Supplementary Table S1 . The sample collection, Enterococcus detection, and identification methods are presented in theSupplementary Material . -
Antibacterial susceptibility to a panel of agents was determined by broth microdilution and interpreted according to the Clinical & Laboratory Standards Institute (CLSI) interpretive criteria. The minimum inhibitory concentration (MIC) of 10 antibacterial compounds was tested: ampicillin, penicillin, erythromycin, ciprofloxacin, daptomycin, vancomycin, tetracycline, chloramphenicol, high-level gentamicin (HLGA), and high-level streptomycin (HLSA). An isolate was defined as MDR if it exhibited resistance to 3 or more antibacterial compounds of different classes.
-
Standard E. faecium (9) and E. faecalis (10) MLST schemes were performed. The CCs of E. faecium and E. faecalis were annotated as previously described (7).
-
Statistical analysis was performed using SPSS Statistics for Windows, version 17.0 (SPSS Inc., Chicago, IL, USA). Categorical variables were compared using Pearson’s chi-squared and Fisher’s exact tests. Statistical significance was set at P<0.05.
-
As shown in
Supplementary Figure S1A andSupplementary Table S1 , most samples (488/694, 70.3%) were positive for enterococci. Enterococci were isolated from most human (65/87, 74.7%) and pig (184/224, 82.1%) samples. The prevalence of enterococci in urban environmental samples (8/47, 17.0%) was significantly lower than in other sample categories. Only 50.0% (20/40) of the fruit samples were positive for enterococci, significantly lower than the vegetable (32/37, 86.5%) and salad (19/23, 82.6%) samples.Supplementary Figure S1B andSupplementary Table S2 show the distribution of Enterococcus species across the various sample sources. Eight Enterococcus species were identified among the 488 isolates. The most common species was E. faecalis (358/488, 73.4%), followed by E. faecium (69/488, 14.1%) and E. casseliflavus (26/488, 5.3%).The detection rate of E. faecalis was significantly higher than that of E. faecium in all sample categories (
Supplementary Figure S1C ). E. casseliflavus comprised 45.0% (9/20) of fruit enterococci, while E. faecium comprised 57.9% (11/19) of salad enterococci. -
The resistance levels against a panel of 10 antibacterial compounds are shown in Figure 1A and
Supplementary Table S3 . Resistance to tetracycline (78.3%) and erythromycin (75.4%) was common, while low resistance levels were noted for daptomycin (1.6%) and vancomycin (1.6%). A proportion of Enterococcus isolates (67.0%) were defined as MDR bacteria. E. faecium resistance to ampicillin and penicillin was significantly higher than that of E. faecalis.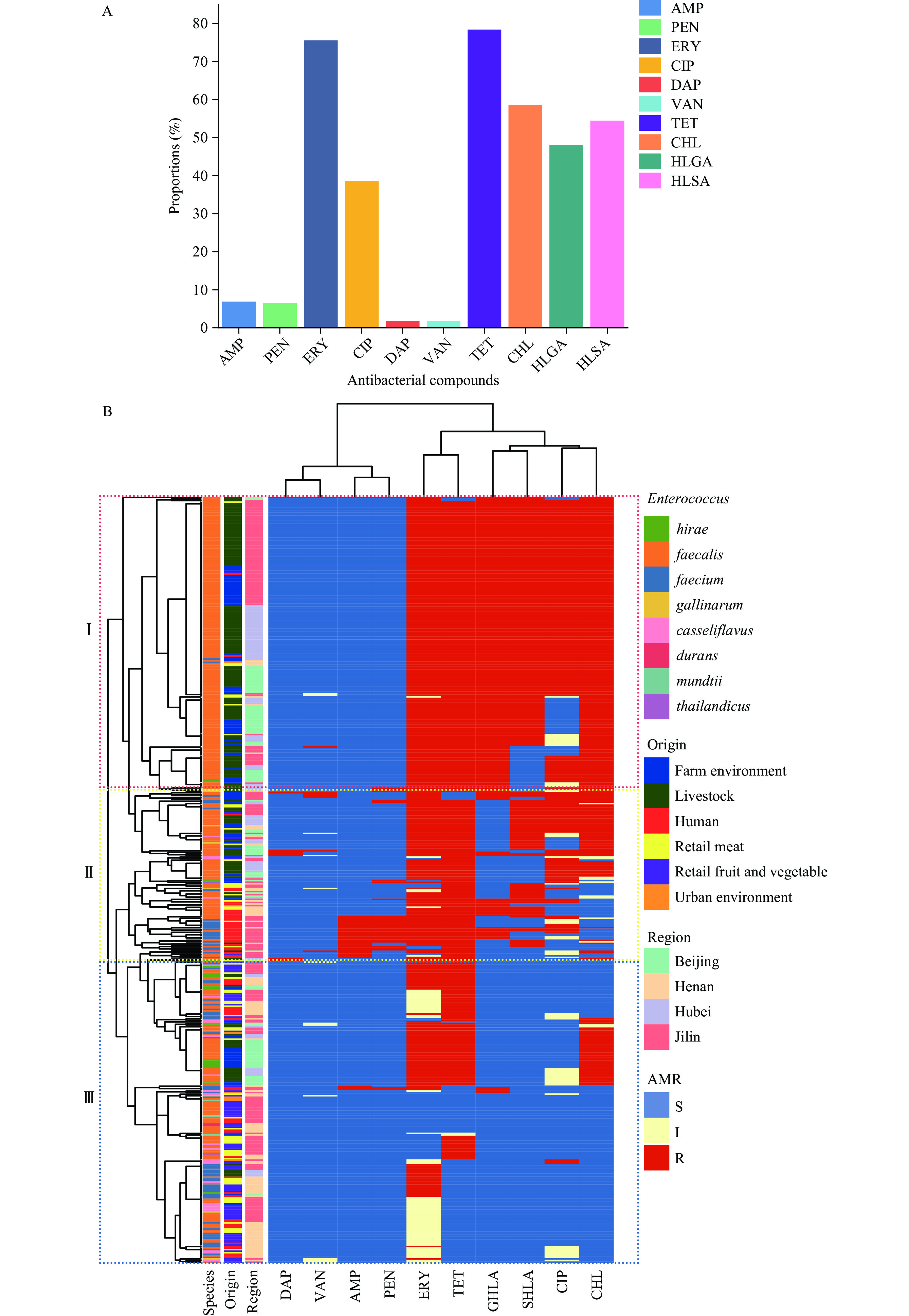 Figure 1.
Figure 1.The enterococcal resistance levels against a panel of (A) ten antibacterial compounds; (B) hierarchical clustering and antibacterial resistance (ABR) heat map of enterococcal isolates from various sampling sources.
Note: (B) Part I shows that E. faecalis from livestock and the farm environment was the least susceptible to antibiotics; (B) Part II shows that multiple Enterococcus species were present in humans and retail meat samples; (B) Part III shows that Enterococcus isolates from retail fruits and vegetables and urban environments were the most susceptible to the panel of antibacterial compounds tested. Antibacterial susceptibility clusters were constructed using the hclust package in R with complete linkage as the default (https://www.rdocumentation.org/packages/stats/versions/3.6.2/topics/hclust). The ABR heatmaps were drawn using the pheatmap package in R (https://www.rdocumentation.org/packages/pheatmap/versions/1.0.12/topics/pheatmap).
Abbreviation: AMP=ampicillin; PEN=penicillin; ERY=erythromycin; CIP=ciprofloxacin; DAP=daptomycin; VAN=vancomycin; TET=tetracycline; CHL=chloramphenicol; HLGA=high-level gentamicin; HLSA=high-level streptomycin; S=susceptible; I=intermediate; R=resistant.
E. faecalis from livestock and the farm environment exhibited the lowest susceptibility to antibiotics, followed by multiple Enterococcus species from humans and retail meat. Isolates from retail fruits and vegetables and urban environments were the most susceptible to the tested panel of antibacterial compounds (Figure 1B).
A comparison of ABR enterococcal isolates recovered from humans, food, farms, and the environment is shown in Figure 2A. Approximately half (50.8%, 33/65) of the human Enterococcus isolates were MDR. Enterococcal isolates recovered from the diarrheal feces of children were MDR, including resistance to ampicillin and penicillin, unlike those retrieved from other human sample types (Figure 2B). Furthermore, human E. faecium was more prone to express an MDR phenotype than E. faecalis.
 Figure 2.
Figure 2.Comparison of the resistance levels of enterococci recovered from human, food, farm, and environmental sources against (A) a panel of ten antibacterial compounds. Hierarchical clustering and antibacterial resistance heat map of enterococcal isolates recovered from (B) human, (C) food, (D) farm, (E) and environmental sources.
Note: Enterococcal isolates recovered from the diarrheal feces of children were multidrug-resistant, unlike those retrieved from other human sample types (B, Part Ⅰ and Ⅱ). Most E. faecalis isolates from animal food samples (chicken, pork, and beef) were resistant to ≥5 antibiotics (C, Part Ⅰ); most isolates from vegetables, cooked meat, and salads were sensitive to nearly all tested antibiotics (C, Part Ⅱ); most isolates from Jilin Province were E. faecalis and resistant to tetracycline (TET; C, Part Ⅲ). In farm samples (D, Part Ⅰ), some E. faecium isolates recovered from pig nasal passages, and E. hirae isolated from pig feces were resistant to only one or two antibiotics (ERY or/and TET). Urban environmental and soil enterococcal strains showed less antibiotic resistance than those recovered from the pig environment (E, Part I); E. hirae recovered from the pig barn environment had the resistance profile of TET-CHL-ERY (E, Part Ⅱ).
Abbreviation: AMP=ampicillin; PEN=penicillin; ERY=erythromycin; CIP=ciprofloxacin; DAP=daptomycin; VAN=vancomycin; TET=tetracycline; CHL=chloramphenicol; HLGA=high-level gentamicin; HLSA=high-level streptomycin; S=susceptible; I=intermediate; R=resistant.
Food (meat, fruits, vegetables, and salads) enterococci exhibited significantly lower resistance to ciprofloxacin (10.5%), HLGA (10.5%), and HLSA (27.8%) than enterococci from other sources. The MDR rate in food enterococci was 20.3% (27/133). Most isolates resistant to ≥5 antibiotics were E. faecalis. Most isolates from vegetables, cooked meat, and salads were susceptible to nearly all tested antibiotics (Figure 2C).
Farm enterococci exhibited significantly higher resistance to erythromycin (98.6%) than enterococci from other sources (Figure 2D). Nearly all (93.6%, 264/282) Enterococcus isolates from farms were MDR-positive, and most were resistant to four or five antibiotics. However, some E. faecium isolated from pig nasal passages, and E. hirae isolated from pig feces, were resistant to only one or two antibiotics (erythromycin or tetracycline).
Environmental enterococci exhibited significantly higher resistance to tetracycline, chloramphenicol, HLGA, and HLSA than those isolated from human and food sources. A large proportion (87.7%, 93/106) of environmental enterococci were MDR. Urban environmental and soil enterococcal strains exhibited less antibiotic resistance than pig environmental strains (Figure 2E). E. hirae isolated from the pig barn environment was resistant to tetracycline, chloramphenicol, and erythromycin.
-
E. faecium exhibited significantly higher resistance to ampicillin and penicillin than E. faecalis (Figure 3A). The MDR rates among E. faecalis and E. faecium isolates were 78.2% (280/358) and 36.2% (25/69), respectively. Most MDR E. faecalis isolates were from livestock and the farm environment. In contrast, most E. faecalis isolates of human and food origin were non-MDR strains (Figure 3B). This finding contrasted with E. faecium, in which most isolates from humans were defined as MDR, while most isolates from food and livestock were resistant to no more than two antibiotics (Figure 3C).
 Figure 3.
Figure 3.Comparison of E. faecalis and E. faecium resistance levels against (A) a panel of ten antibacterial compounds. (B) Hierarchical clustering and antibacterial resistance heat maps of E. faecalis and (C) E. faecium isolated from all tested samples.
Note: Most E. faecalis isolates of human and food origin were non-MDR strains (B, Part I). Most isolates from food and livestock were resistant to no more than two antibiotics (C, Part I).
Abbreviation: AMP=ampicillin; PEN=penicillin; ERY=erythromycin; CIP=ciprofloxacin; DAP=daptomycin; VAN=vancomycin; TET=tetracycline; CHL=chloramphenicol; HLGA=high-level gentamicin; HLSA=high-level streptomycin; S=susceptible; I=intermediate; R=resistant.
-
The E. faecalis isolates showed highly heterologous genotypes, with 95 STs among the 358 E. faecalis isolates, including 28 (29.5%) new STs. The main E. faecalis STs were ST4 (10.3%), ST86 (7.5%), ST476 (6.2%), and ST330 (5.9%; Figure 4A). Two E. faecalis CCs, CC16 (major in ST16) and CC21 (major in ST21), were identified. Three major CC and ST clades (CC21, CC16, and ST69-ST632), covering the isolates from children’s diarrheal feces, pigs, meat, vegetables, and fruits, E. faecalis ST16 and ST65 had complex sample sources, including animals, food, and patients’ diarrheal feces. As shown in Figure 4B, the CC16 clade, ST4, ST16, ST86, ST330, and other isolates were identified as MDR.
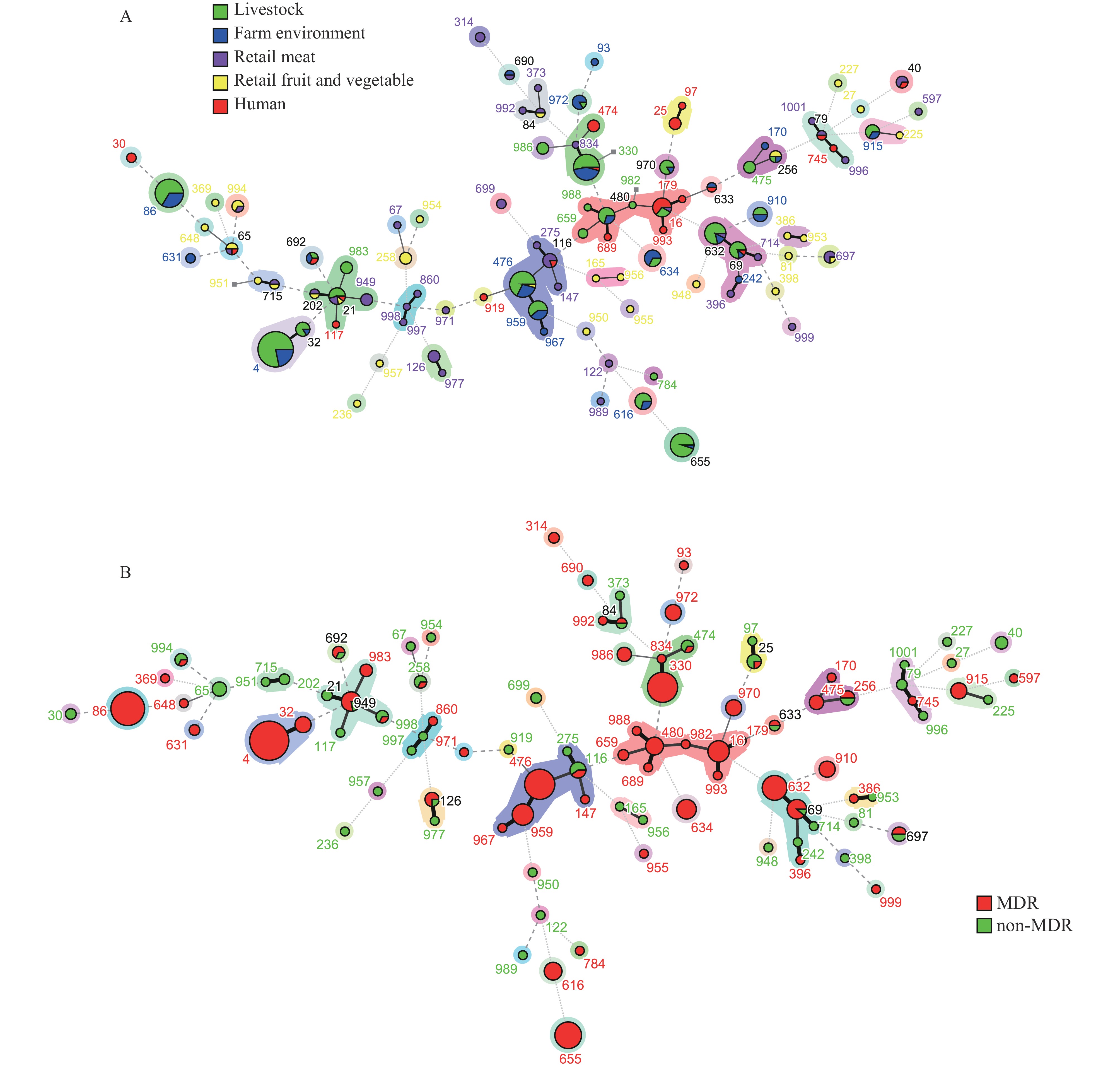 Figure 4.
Figure 4.A minimum spanning tree of (A) E. faecalis STs, (B) and MDR isolate distribution as determined by multilocus sequence typing.
Note: The node size reflects the number of isolates included in that particular clade.
Abbreviation: ST=sequence type; CC=clonal complex; MDR=multidrug resistance.
The 69 E. faecium isolates analyzed were divided into 44 STs, including 21 (47.7%) novel STs. The main E. faecium STs included ST94 (11.6%), ST569 (8.7%), and ST296 (7.3%) (Figure 5A). Unlike the E. faecalis isolates, which exhibited complex prevalent ST and clade structures, the E. faecium isolates were primarily divided into two distinct CC clades, CC17 and CC94. The main isolates in the CC17 clade were recovered from children’s diarrheal feces, while CC94 isolates were mainly from retail fruits and vegetables, and pigs. Most isolates in the CC17 clade were MDR, while most isolates in the CC94 clade were non-MDR (Figure 5B). ST60 and ST94 isolates originated from children’s diarrheal feces and ready-to-eat food. Notably, despite belonging to the same STs, isolates from clinical patients exhibited MDR phenotypes, while isolates from food were non-MDR.
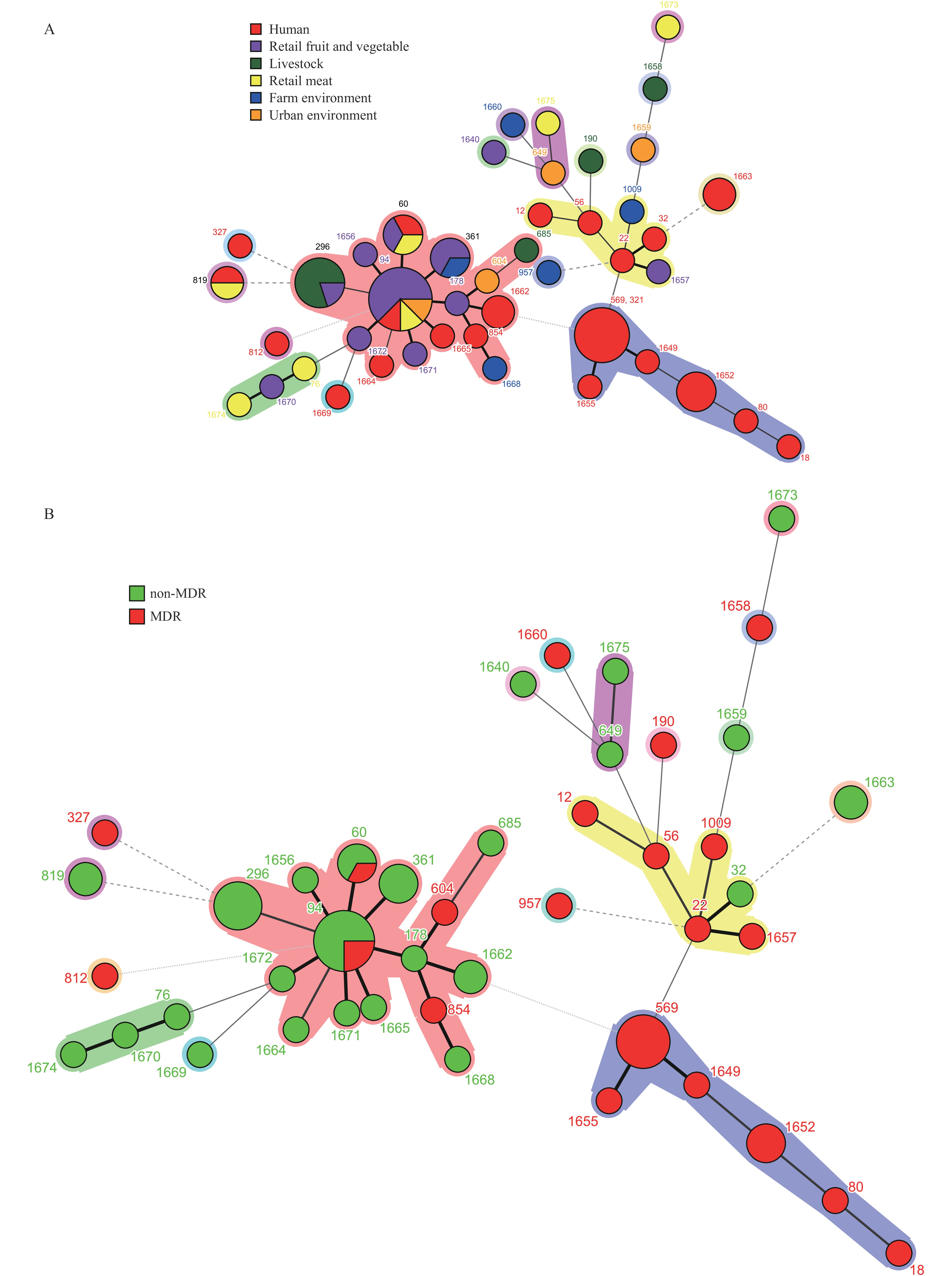 Figure 5.
Figure 5.Minimum spanning tree of (A) E. faecium STs, (B) multidrug-resistant isolate distribution as determined by multilocus sequence typing.Abbreviation: ST=sequence type; CC=clonal complex.
-
Although enterococci are considered opportunistic pathogens, they can be reservoirs of ABR and transfer resistance to humans through the food chain (11). This study demonstrated that enterococci were abundant across most parts of the food chain in China, particularly in animal farms and humans. The most dominant species identified was E. faecalis, followed by E. faecium and E. casseliflavus. E. faecium CC17 isolates were dominant in children’s diarrheal feces, whereas E. casseliflavus was the dominant species in retail vegetables and fruits.
A high level of MDR enterococci was found among food-producing animals and the animal farm environment, possibly because antibiotics are widely used as animal growth promoters in China. Enterococci in commercial food animal production and human feces could contaminate the food chain during processing or through the composting of these wastes for use as biofertilizers on farms (12). It has been reported that many E. faecium bloodstream infections were of gastrointestinal origin, raising the possibility that food might be a vehicle for such bacteria (13). Our data showed that farms and foods were highly contaminated by enterococci and that the most dominant species was E. faecalis, followed by E. faecium. Farm, food, and environmental enterococci exhibited high resistance to tetracycline, erythromycin, and HLSA. Notably, E. faecalis ST16 and ST65 and E. faecium ST60 and ST94 were found in isolates from farms, food, and patients’ diarrheal feces, suggesting potential pathogen transfer along the food chain.
To treat enterococcal infections, first-choice antibiotics are typically β-lactam-based compounds and aminoglycosides. Second-choice antibiotics include glycopeptides, especially vancomycin (14). However, in this study, most enterococci isolated from children’s diarrheal feces belonged to the E. faecium hospital-lineage CC17 clade, exhibiting high resistance to ampicillin, penicillin, HLGA, and HLSA, which limits treatment options. The E. faecium CC94 clade, a traditional community lineage, was detected in diarrheal feces from patients, suggesting that community isolates may be spreading into hospitals. Interestingly, the hospital CC94 isolates, ST60 and ST94, were MDR, whereas community isolates with the same ST were not. This suggests that hospital environments, with their greater use of antibacterials and disinfectants, may promote MDR formation in hospital-adapted isolates compared to community-acquired isolates. However, sampling sources were imbalanced across regions. No inpatient samples were collected in Beijing Municipality and Hubei Province due to ethical concerns, affecting the completeness of the One Health approach in this study.
In conclusion, our results suggest that effective precautionary measures should be taken to prevent the occurrence of MDR enterococci in the food chain.
-
Tania Dottorini, Michelle Baker, and Necati Esener of the University of Nottingham for helping with data analysis and visualization.
HTML
Sample Collection and Enterococcus Detection and Identification
Antibacterial Susceptibility Testing
E. faecium and E. faecalis Multilocus ST (MLST) and CC Data Analysis
Statistical Analysis
Prevalence of Enterococci and Species Diversity in the Food Chain
ABR of Enterococcus Species
ABR of E. faecalis and E. faecium
STs and CCs of E. faecalis and E. faecium
| Citation: |

|