

 Download:
Download:




-
The emergence of Bartonella species has been identified as a cause of blood-culture-negative endocarditis. While the list of Bartonella species carried by rodents has rapidly expanded, species carried by Marmota himalayana (M. himalayana) remain unknown. To investigate this, Bartonella washoensis (B. washoensis) screening was conducted using samples obtained from both deceased and captured marmots during plague surveillance in Jiuquan City, Gansu Province from 2021 to 2022. The B. washoensis species were identified through 16s rRNA gene and multi-locus sequence typing (MLST), with phylogenetic trees constructed using the neighbor-joining method.
The detection rate of B. washoensis in captured marmots (29.58%, 21/71) was found to be significantly higher than that of deceased marmots (10.28%, 11/107), with relatively high rates observed in marmot heart and lung samples. The new sequence type, Sequence Type 22 (ST22), discovered in marmots, possessed five loci of novel sequences and clustered between B. washoensis from Spermophilus dauricus in China and Spermophilus columbianus in the United States. Importantly, human endocarditis-associated B. washoensis was identified for the first time and demonstrated a high prevalence in M. himalayana.
Our findings suggest that B. washoensis in marmots may have a preference for heart and lung tissue, and individuals in specific areas may be at risk of B. washoensis infection. Consequently, there is a pressing need for continued surveillance of B. washoensis and identification of clinical cases.
Overall, 13 of the 45 known Bartonella species are documented to infect humans. Species such as Bartonella quintana (B. quintana), Bartonella henselae (B. henselae), B. washoensis, Bartonella koehlerae (B. koehlerae), Bartonella alsatica, Bartonella elizabethae, and others have emerged as causes of blood-culture-negative endocarditis (1). Among these species, B. henselae and B. quintana are the most frequent causes of infectious endocarditis in humans (2-3). Recently, the list of Bartonella species carried by rodents has rapidly expanded. M. himalayana is the primary host of Yersinia pestis (Y. pestis) and is also known to carry a variety of bacteria and parasites pathogenic to humans (4-5). Although Bartonella has been reported in M. himalayana in China, the specific infecting species of Bartonella remains unidentified (6). In this study, human endocarditis-associated B. washoensis was initially identified in M. himalayana during the plague surveillance conducted from 2021 to 2022, and its public health risks were analyzed.
Peripheral blood mononuclear cells (PBMCs) were isolated from M. himalayana blood samples collected during plague surveillance in the Altun Mountains, which is part of the M. himalayana plague focus in the Qinghai-Tibet Plateau (7). DNA was extracted from the samples using the DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany). The 16S rRNA gene (27F, 1492R) was amplified, sequenced, and aligned using BLASTn. Additionally, six housekeeping loci of B. washoensis (16S rRNA gene, ftsZ, gltA, groEL, ribC, and rpoB) were amplified and sequenced (8). Allele numbers and sequence type (ST) assignments were determined through pubMLST (9).
Sequences were extracted from the reference genome of 40 Bartonella species by aligning with six housekeeping genes from B. washoensis. A phylogenetic tree was constructed using the neighbor-joining (NJ) method based on the concatenated sequence of these six housekeeping genes. NJ trees were generated for both the Bartonella genus and the B. washoensis species. In the B. washoensis species tree, four Bartonella species closely related to B. washoensis (B. quintana, Bartonella senegalensis, B. henselae, and B. koehlerae) were also incorporated.
During the 2021–2022 plague surveillance in the Altun Mountains, samples were collected from six organs (heart, liver, spleen, lung, kidney, and bone) of 107 M. himalayana found dead in the environment and 71 live-captured marmots. These findings were within the M. himalayana plague focus of Qinghai-Tibet Plateau. The B. washoensis gltA gene was screened, sequenced, and aligned according to a previously published method (10). Detection rates were compared between deceased and live marmots, among the six sampled organs, and between B. washoensis and Y. pestis.
Chi-squared tests were used to compare the differences between groups, with P-values<0.05 considered statistically significant. Fisher’s exact test was employed if the theoretical frequency ranged between 1<T<5. Statistical analyses were conducted using the SPSS software (version 19.0, IBM Corp., NY, USA).
The highest BLASTn match for the 16S rRNA gene from marmots in our study was B. washoensis (GenBank: AB519060.1), exhibiting 100% coverage and 100% identity. Other Bartonella species, such as Bartonella volans (GenBank: EU294521.1), were identified with 100% coverage and 99.77% identity, surpassing the species identification threshold (10). Based on the NJ tree of 40 Bartonella species, which was constructed using concatenated sequences of housekeeping genes, the sequences derived from marmots formed a cluster on the same branch as B. washoensis. Neighboring branches included B. quintana, B. senegalensis, B. henselae, and B. koehlerae (
Supplementary Figure S1 ).The housekeeping gene allele numbers of B. washoensis in marmots were identified as 2-15-18-20-20-18 for the 16S rRNA gene, ftsZ, gltA, groEL, ribC, and rpoB, which collectively formed the novel ST 22. Excluding the 16S rRNA gene, the other five genes displayed at least 5, 8, 18, 14, and 26 base differences from known sequences. The NJ trees for B. washoensis (Figure 1) revealed that B. washoensis isolates from marmots clustered together with those from ground squirrels (Spermophilus), showing the greatest similarity to B. washoensis from S. columbianus. The other four Bartonella species (B. quintana, B. senegalensis, B. henselae, and B. koehlerae) formed separate clusters outside the B. washoensis strains.
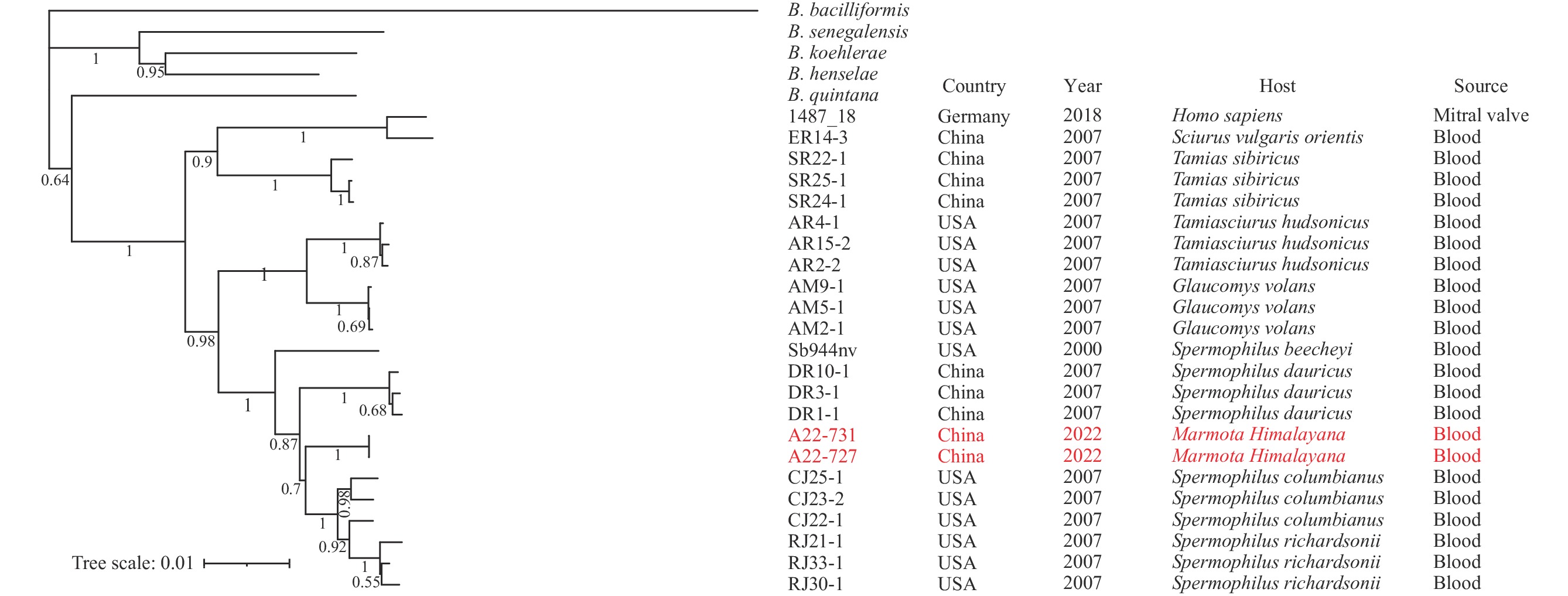 Figure 1.
Figure 1.Phylogenetic trees of B. washoensis based on concatenated sequences of housekeeping genes. Note: Red means in this study.
The detection rate of B. washoensis in captured marmots (29.58%, 21/71) was found to be significantly higher than that in self-dead marmots (10.28%, 11/107, χ2=10.778; P=0.001). In the captured marmots, the top four positive organs were bone (29.27%, 12/41), spleen (15.52%, 9/58), lung (13.56%, 8/59), and heart (9.30%, 4/43) (Table 1). Statistical differences were observed between the highest rate in the bone (29.27%) and the lowest in the liver (1.72%, χ2=19.159; P<0.002). For self-dead marmots, the top four positive organs included spleen (17.50%, 7/40), lung (9.76%, 4/41), heart (7.69%, 3/39), and bone (4.00%, 4/100), with no significant differences between different organs (Fisher exact test, χ2=9.573; P=0.059). The detection rate was also lowest in the liver (2.38%, 1/42) in self-dead marmots. Moreover, B. washoensis positive rates showed no differences between marmots with or without Y. pestis (
Supplementary Table S1 ) (χ2=0.628; P=0.428). B. washoensis was detected in 7.84% (4/51) of Y. pestis-negative self-dead marmots, and in 12.5% (7/56) of Y. pestis-positive self-dead marmots. The 77 B. washoensis-positive samples revealed three different gltA sequence types, displaying one to three single nucleotide polymorphisms (SNPs), all of which were synonymous mutations.Sample Captured marmot Self-dead marmot Positive Total Positive rate (%) Positive Total Positive rate (%) Bone 12 41 29.27* 4 100 4.00 Spleen 9 58 15.52 7 40 17.50 Lung 8 59 13.56 4 41 9.76 Heart 4 43 9.30 3 39 7.69 Kidney 3 43 6.98 1 37 2.70 Liver 1 58 1.72* 1 42 2.38 * means statistical significance between positive rates of bone and liver in captured marmots. Table 1. Detection rates of B. washoensis in six organs of marmots.
-
In the current study, the first identification of human endocarditis-associated B. washoensis was observed with a relatively high detection rate in M. himalayana (approximately 1/3 in captured marmots and 1/10 in self-dead marmots). An increasing number of Bartonella species have been found to contribute to blood culture-negative endocarditis, with B. henselae and B. quintana being the most common causes of human cases (2-3) (
Supplementary Figure S1 ) (double asterisk). B. washoensis has recently been associated with human and dog endocarditis (10-11) and exhibits a close relationship with B. quintana and B. henselae in the NJ tree, based on housekeeping gene sequences. B. washoensis was detected in six types of marmot organs, suggesting a systemic distribution in its carriers. The spleen, heart, lung, and bone displayed the highest detection rates. The order of rates in captured and self-dead marmots varies, which may be attributable to sample freshness or other causes of death. Nonetheless, higher rates were detected in the heart and lung of marmots, implying that B. washoensis may exhibit a preference for these organs. In light of its phylogenetic position in the NJ tree, it is suggested that B. washoensis from M. himalayana closely resembles human-endocarditis-related Bartonella. Moreover, the presence of Y. pestis does not impact B. washoensis, and B. washoensis may serve as a cause of marmot mortality independent of Y. pestis. Lastly, the detection rate of B. washoensis is lowest in the liver of marmots, rendering the spleen the recommended site for detection.The detection of B. washoensis in M. himalayana has expanded the known range of rodent species that carry this bacterium, while the discovery of the novel ST22 has enhanced its genetic lineage by adding a marmot-origin branch between S. columbianus and S. dauricus. B. washoensis has been previously identified in squirrels from the provinces of Hebei and Zhejiang in China (12). However, the housekeeping gene sequences of B. washoensis in marmots and squirrels exhibit significant differences. Five of the six housekeeping genes, excluding the 16S rRNA gene, have 5–26 SNPs when compared with the closest known sequences. Most gltA genes of B. washoensis observed in marmots differ by two bases, suggesting their close relationship with the ST22 type. The geographical isolation of the Qinghai-Tibet Plateau may contribute to the divergence of pathogens in M. himalayana, such as the new bacterial species Streptococcus respiraculi (13), Helicobacter himalayensis (14), and the novel parasite species Enterocytozoon bieneusi (5). Our research team will continue efforts to isolate and purify B. washoensis from marmots in order to further elucidate its biological and genomic characteristics.
In this study, we identified human endocarditis-associated B. washoensis for the first time, with a relatively high detection rate in M. himalayana and even higher in the heart and lungs of marmots. The new sequence type, ST22, of M. himalayana-derived B. washoensis is considerably different from the previously reported housekeeping gene sequence, expanding the isolation source and genetic lineage of B. washoensis. Bartonella has become an emerging cause of human endocarditis worldwide; however, related monitoring, detection, and diagnosis are insufficient in our country. Our findings suggest that individuals in high-risk areas are susceptible to B. washoensis infection, necessitating public health warnings and enhanced clinical case identification.
-
No conflicts of interest.
-
Charlesworth Author Services (Paper no.116713) for their critical editing and helpful comments regarding our manuscript.
HTML
| Citation: |

|