

 Download:
Download:




-
The ongoing severe acute respiratory syndrome coronavirus type 2 (SARS-CoV-2) pandemic, with the increasing number of coronavirus disease 2019 (COVID-19) cases, poses a severe challenge to public health (1). The antigen-specific adaptive immunity in COVID-19 convalescents and vaccinated population is crucial for long-term protection upon individual exposure to SARS-CoV-2 (2-3). In addition to humoral immunity, which produces antibodies to neutralize the virus, T cells play an indispensable role in the process of virus clearance (4). CD8+ T cells recognize antigenic peptides presented by major histocompatibility complex I (MHC-I) through T cell receptors (TCRs) and are activated to kill virus-infected target cells (4-5). Nucleocapsid (N) protein is one of the main structural proteins of SARS-CoV-2 and can trigger broad T-cell responses in humans (6). Herein, an overlapping peptide pool covering the full length of N protein was designed, overlapping peptides with positive T-cell activating potency in COVID-19 convalescents were screened, and CD8+ T cell epitopes in the positive overlapping peptides were further identified. Through enzyme-linked immunospot assay (ELISpot), a human leukocyte antigen A (HLA-A)*1101-restricted CD8+ T cell epitope, which could stimulate the production of IFN-γ by the peripheral blood mononuclear cells (PBMCs) of the convalescents, was defined, and the tetramer generated with this epitope could detect SARS-CoV-2-specific T cells in the PBMCs of the convalescents. The in vitro refolding experiments and crystallographic structural analysis showed that the epitope was a typical HLA-A*1101-restricted and conserved among all the sarbecoviruses. These findings provided the basis for cellular immunity evaluation in COVID-19 recovered patients and vaccinated donors, and the newly identified epitope was also helpful for the development of a polypeptide vaccine for SARS-CoV-2.
-
A total of 5 convalescent patients in Macheng City, Hubei Province, who had recovered from COVID-19 for 6 months, were enrolled and their anticoagulant blood was collected to isolate PBMCs. At the same time, the DNA of the PBMCs was extracted for HLA type sequencing. Peptides with the length of 15- to 18-mer overlapping by 10 amino acids derived from the SARS-CoV-2 N protein (NCBI Reference Sequence: YP_009724393.1) were designed using the PeptGen website (
https://hcv.lanl.gov/content/sequence/PEPTGEN/peptgen.html ). Potential CD8+ T cell epitopes within the 15- to 18-mer peptides were predicted by NetMHC website (https://services.healthtech.dtu.dk/service.php?NetMHC-4.0 ). Peptides with a purity of over 95% were synthesized by Beijing Scilight Biotechnology Corporation, and the peptide powders were dissolved in DMSO before use. An HLA-A*1101 tetramer complexed with SARS-CoV-2-specific peptide was generated in our laboratory as described previously for the preparation of other HLA class I tetramers (6).For in vitro PBMCs culture, the N peptide pools (2 μg/mL single peptide), recombinant IL-7 (20 μg/mL, Pepro Tech, USA), and recombinant IL-2 (0.02 μg/mL, Pepro Tech, USA) were added to PBMCs that had been cultured in a 24-well Costar plate. Half of the cultured medium was replaced every three days. After nine days of incubation, the cells were harvested for ELISpot, intracellular cytokine staining (ICS) and flow cytometry-based tetramer staining as described previously (7). HLA-A*1101 molecules were expressed through prokaryotic inclusion bodies and refolded with the epitope by gradual dilution methods in vitro. After the complex was purified by Superdex 200 Increase 10/300 GL chromatography column (GE Healthcare, China), the crystal growth conditions were screened using the sitting-drop vapor diffusion technique and X-ray diffraction was used to distinguish the quality of the collected protein crystal. Flowjo (version10, BD, New Jersey, USA) was used for flow cytometry analysis, Origin (version 9.6.5, OriginLab, Massachusetts, USA) was used for refolding result analysis, and PyMOL (version 2.3.2, Schrodinger, New York, USA) was used for crystal structure analysis.
-
We obtained anticoagulant blood samples from 5 convalescent patients (Supplementary Table S1) with different HLA haplotypes for peptides screening. A total of 57 overlapping peptides of SARS-CoV-2 N protein were designed in this study. ELISpot experiment was performed with PBMCs of convalescent patients under the stimulation of different overlapping peptides. Within the positive overlapping peptides that could stimulate T cells to secrete IFN-γ, peptide nCoV-N49 showed the highest response intensity. Subsequently, potential CD8+ T cell epitopes from nCoV-N49 were predicted according to HLA haplotype of the positive convalescent patient (Table 1). The epitope named N25 (KTFPPTEPK) with HLA-A*1101 restriction was verified as positive by ELISpot.
Peptide nCoV-N49 Predicted epitope Position HLA typing Affinity (nmol/L) Affinity (nmol/L) LLNKHIDAYKTFPPTEPK KTFPPTEPK† 361–369 A*1101 6.28 0.01 LLNKHIDAYK 352–361 A*1101 149.83 0.9 YKTFPPTEPK 360–369 A*1101 19.13 0.1 Abbreviations: SARS-CoV-2=severe acute respiratory syndrome coronavirus type 2; HLA=human leukocyte antigen.
† The peptide was named N25.Table 1. Predicted CD8+ T cell epitopes derived from long peptide nCoV-N49 of SARS-CoV-2.
We selected one HLA-A*1101-positive convalescent patient (ID:11) for intracellular cytokine staining based on flow cytometry. The in vitro cultured PBMCs were harvested; after being stimulated with N25 epitope for 10 h, the PBMCs were stained with fluorescent antibodies. The result showed that PBMCs from the convalescent donor could secrete detectable IFN-γ after being stimulated by N25 epitope. The proportion of IFN-γ-positive CD8+ T cells was 1.30%, which was higher than 0.22% of the control sample without epitope stimulation (Figure 1A). Then the cultured PBMCs with the HLA-A*1101/N25 tetramer were stained and analyzed by flow cytometry, immediately. The N25 tetramer staining showed 3.59% of the CD8+ T cells were specific for peptide N25 in the convalescent patient, and the control tetramer was detected as 0.24% (Figure 1B).
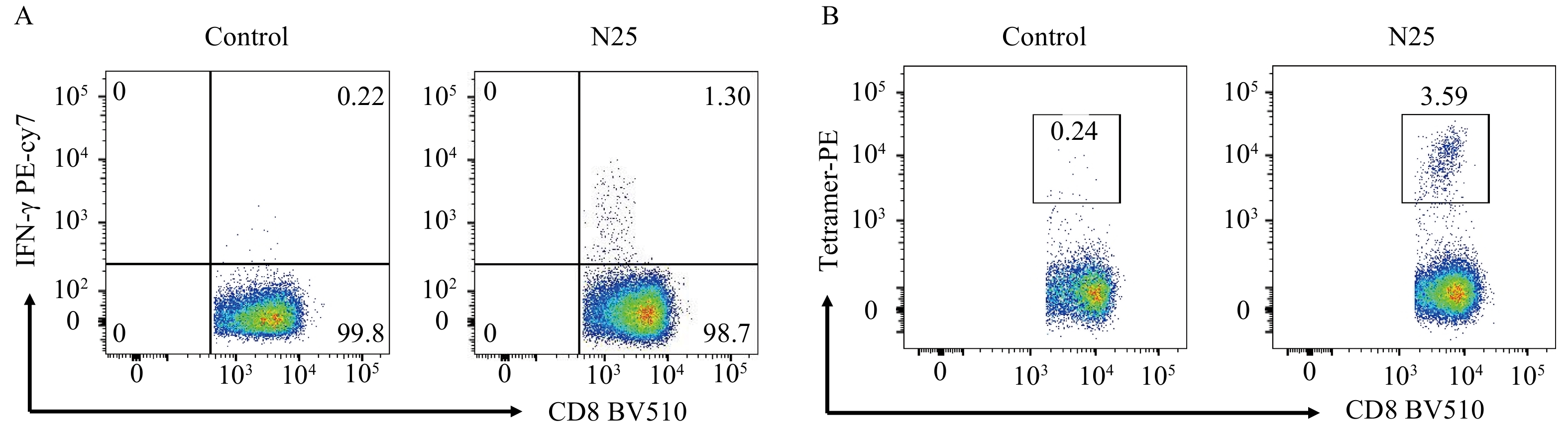 Figure 1.
Figure 1.The SARS-CoV-2-specific CD8+ T cells detection in COVID-19 convalescents using the peptide-based IFN-γ-secreting intracellular cytokine staining and tetramer staining in flow cytometry. (A) Intracellular cytokine (IFN-γ) staining, negative control (left), positive sample (right); (B) Detection of SARS-CoV-2-specific CD8+ T cells using N25/HLA- A*1101 tetramer.Abbreviations: SARS-CoV-2=severe acute respiratory syndrome coronavirus type 2; COVID-19=coronavirus disease 2019; HLA=human leukocyte antigen.
To confirm the binding feature of peptide N25 with HLA-A*1101, in vitro folding assay was conducted. The heavy and light chains of HLA-A*1101 molecule were expressed in the form of inclusion bodies by the expression system of Escherichia coli. Then the heavy and light chain proteins were renatured with epitope N25 in vitro. The results showed peptide N25 could help to form the stable HLA-A*1101 complex under the assistance of N25 and that the purified complex eluted at 16.5 mL during purification by chromatography column. The purified complex had the highest peak that could represent efficient renaturation. The formation of the HLA-A*1101/N25 complex was also verified by identification of two protein bends: HLA-A*1101 and β2 microglobulin in SDS-PAGE (Figure 2A).
 Figure 2.
Figure 2.The HLA-A*1101-binding of newly identified SARS-CoV-2 T-cell epitope and the structural conformation of the peptide presented by HLA-A*1101. (A) The abilities of peptide N25 presented by HLA-A*1101 were evaluated using in vitro refolding and SDS-PAGE. M: protein ladder; P1: Peak 1, presents the heavy chain aggregates; P2: Peak 2, presents the N25/HLA-A*1101 complex; P3: Peak 3, presents the light chain. (B) Crystal of N25/HLA-A*1101 complex. (C) The overall conformation of the structurally-defined peptide N25 was shown with green sticks. The carbon, nitrogen, and oxygen atoms were shown in green, blue, and red, respectively. The α1 helix and β-sheets of HLA-A*1101 were shown in grey stripes. (D) Alignment of the SARS-CoV-2 N25 peptide amino acid sequence with other sarbecoviruses.Abbreviations: SARS-CoV-2=severe acute respiratory syndrome coronavirus type 2; HLA=human leukocyte antigen; SDS-PAGE=Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis.
To show whether N25 was a HLA-A*1101-restricted T-cell epitope with the classic peptide conformation, the crystal structure of HLA-A*1101/N25 complex was determined. The HLA-A*1101/N25 complex protein was purified, and then used to screen the protein crystals by the crystal screening kit (Crystal Screen, Hampton Research, California, USA) at a concentration of 15 mg/mL. On Day 7, the HLA-A*1101/N25 complex was observed to grow one rhombus crystal at 18 ℃ using the buffer solution 2.0 M Ammonium sulfate of CS I kit (Figure 2B). The crystal was sent to Shanghai Synchrotron Radiation Facility and the structure was determined by X-ray crystallography to 1.5 Å resolution (Supplementary Table S2). As other HLA-A*1101-presented peptides, the residues at position 2 (P2) and C-terminus (PΩ) of peptide N25 act as anchor residues (8-9). The ability of MHC to bind peptides is mainly determined by the interaction between the residues at P2 and PΩ of peptide with the B and F pocket of MHC molecule, respectively (10). According to our previous studies, the HLA-A*1101 molecule preferred serine, threonine, or aliphatic amino acids at the second residue of peptide, while the PΩ of HLA-A*1101-binding peptide was usually lysine (11). N25 epitope fits this pattern very well with the P2 residue threonine in pocket B, and the PΩ residue lysine into pocket F, showing a stable bind to the HLA-A*1101 molecule (Figure 2C). In addition to the primary anchors, i.e. threonine at P2 and lysine at the PΩ positions, the P6 residue threonine also acts as a secondary anchor residue for N25, with the side chain inserting into the peptide binding groove of HLA-A*1101. Except these three residues, other residues of N25 are accessible for T-cell recognition.
The sequence conservation of the peptide N25 among the sarbecoviruses such as SARS-CoV and bat coronavirus RaTG13, etc. (Figure 2D) was also analyzed. N25 is completely conserved among sarbecoviruses. This indicated that the peptide N25 that was identified in our study can be used for the evaluation of the T-cell immunity for all these sarbecoviruses and may also act as a candidate for the T cell-based vaccine for these viruses.
-
Up to now, the SARS-CoV-2 pandemic has been going on for almost two years, and there are diverse methods for the assessment of SARS-CoV-2 antibody levels of convalescent patients or vaccinated persons (7), but there were few reports on the assessment of T-cell immune responses. T-cell immunity, especially CD8+ cytotoxic T lymphocyte, plays an important role in the anti-SARS-CoV-2 process (12). When the virus infects the cell, viral proteins are digested by cellular proteases into MHC-presented peptides so that the MHC/peptides can be recognized by T cell receptors and ultimately activate the CD8+ T cells to clear the virus. Therefore, identification of T-cell epitopes is particularly important in antiviral T-cell immunity (5,13-14). In previous studies on T-cell immunity of other coronaviruses, structural proteins of the viruses, such as N protein in addition to S protein, were also identified as immunodominant antigens, which were able to stimulate the T-cell immune response (15). In our recent study on the T-cell immune evaluation of COVID-19 convalescents, we found that N protein could stimulate a wide range of T-cell immune responses and was an ideal T-cell immunogen (7).
Thus, in this study, CD8+ T cell epitopes of N protein based on the PBMCs of COVID-19 convalescents were screened, and one HLA-A*1101-restricted epitope N25 with high reaction intensity was identified. Through ELISpot and intracellular cytokine staining, it was verified that the epitope could stimulate the secretion of cytokine IFN-γ by PBMCs of convalescent patients. In addition, a certain proportion of virus-specific CD8+ T cells were detected by staining the PBMCs of the convalescent patients with the N25 tetramer. Furthermore, through in vitro renaturation test, it was verified that N25 could stably bind to HLA-A*1101 and the crystal structure showed a typical HLA-A*1101-restricted peptide conformation of N25. These data provided a deeper understanding of the SARS-CoV-2-derived epitope-triggered T-cell responses.
In conclusion, N25 was a CD8+ T cell epitope with robust T-cell immunogenicity. This peptide could be used to stimulate the T-cell responses in the recovered or vaccinated persons to assess their immune status. Tetramers prepared with this peptide can be used as a means of detecting virus-specific T cells in the recovered patients or vaccinated persons. However, due to the HLA restriction of peptide presentation, the epitope can only be used for the detection of specific T cells in HLA-A11 population. It still needs further investigations to determine whether the peptide can be used to detect the T-cell responses among the population with a broader A3 HLA supertype, such as HLA-A03, HLA-A33, and HLA-31. The sequence analyses showed that the sequence of the epitope was conserved in a variety of different coronaviruses, which can be used as a candidate for developing a T cell-based universal vaccine.
-
No conflicts of interest.
-
Macheng CDC and all of the participants.
HTML
| Citation: |

|