

 Download:
Download:




-
By 2050, it is projected that water withdrawals for irrigation will increase by approximately 10%. Utilizing reclaimed water (RW) for irrigation may provide effective strategies to alleviate pressure on groundwater resources (1). While numerous sterilization processes exist to eliminate pathogens and antibiotic-resistant bacteria (ARBs) in RW, antibiotic resistance genes (ARGs) possess a range of mechanisms, such as dormancy and mutations in ARBs, that enable their persistence (2). ARGs and ARBs have the potential to be transferred and reaccumulated in various plant tissues, including those that are edible, thereby entering the food chain. This poses a significant health risk, particularly when these plants are consumed raw.
The dissemination of ARGs in soil-plant systems through RW irrigation is influenced by two primary factors: the colonization of ARBs and the horizontal gene transfer (HGT) of ARGs to indigenous bacteria within these systems. Notably, nanoparticles (NPs) possess the capability to not only alter microbial community structures, thereby affecting the colonization of ARBs (3), but also to potentially inhibit the HGT of ARGs (4). Furthermore, NPs can enhance the efficiency of agricultural resource utilization and more effectively manage environmental challenges, paving the way for a forthcoming nano-agriculture revolution (5). Despite these promising attributes, the role of NPs in reducing ARG dissemination within soil-plant systems during RW irrigation has yet to be fully elucidated.
This study sought to examine the effects of CeO2 NP exposure on radish seedlings grown in soil irrigated with RW. Various application methods were employed — adding NPs to reclaimed water, NP seed dressing, and direct soil mixing with NPs. It was found that, generally, radish plant growth can be significantly enhanced by the presence of CeO2 nanomaterials at concentrations up to 100 ppm (5), which was the predetermined upper limit for this investigation. Following 45 days of irrigation with RW, radishes were harvested and assessed.
A comprehensive evaluation was carried out on the abundance of ARGs and bacterial community diversity in the rhizosphere soil, roots, and leaves of the plants. This was achieved through high-throughput quantitative PCR (HT-qPCR) and 16S ribosomal RNA (rRNA) gene amplicon sequencing. This study provides initial insight into the influence and underlying processes by which NPs might modulate the proliferation of ARGs in RW irrigation systems. Furthermore, these findings are intended to lay a theoretical foundation for the development and refinement of nanotechnologies aimed at preventing and mitigating the spread of the agricultural resistome.
HTML
-
Radishes (Raphanus sativus L. var. radculus pers) were cultivated in a plant seedling substrate soil (Nord Agri, China). Four types of treatments were designed, including non-nanoparticle treatment (CK group), reclaimed water added with 100 ppm CeO2 NPs treatment (WT group), seed dressing by 100 ppm CeO2 NPs treatment (SD group) and soil mixed with 100 ppm CeO2 treatment (ST group) (Figure 1). The reclaimed water, sourced from the Futian Wastewater Treatment Plant (WWTP) in Shenzhen, was used to maintain soil moisture levels with applications every two days. After 45 days, radish plants were harvested, including samples from the rhizosphere soil, roots, and leaves. The dried radish tissue samples were then digested using pure HNO3 through a microwave digestion system. Subsequently, total cerium (Ce) content was quantified using inductively coupled plasma-mass spectrometry (ICP-MS).
 Figure 1.
Figure 1.Schematic workflow depicting experimental design, methods, and objective of this study.
Abbreviation: NPs=nanoparticles; HT-qPCR=high-throughput quantitative PCR. -
The radish shoot, root, and rhizosphere soil were individually weighed and placed into PowerBead tubes for DNA extraction, as per the manufacturer’s protocol. We utilized the HT-qPCR method to examine the diversity and quantify the abundance of ARGs and mobile genetic elements (MGEs) (6). A suite of 296 primer pairs was utilized to profile the antibiotic resistome.
We established a cycle threshold (CT) of 31 as the cutoff for detection (7). The relative gene copy number was calculated using the equation: relative gene copy number = 10(31−CT)/(10/3), in which CT represents the results from the quantitative PCR assay. To determine bacterial abundance, we quantified absolute 16S rRNA gene copy numbers using standard curves. Measurements were conducted using a Bio-rad CFX (Biorad, USA) and a Takara Taq PCR kit (Takara, Japan). -
To characterize the structures and compositions of bacterial communities, the V4–V5 hypervariable region of the 16S rRNA gene was amplified using primers 515F (GTGCCAGCMGCCGCGG) and 907R (CCGTCAATCMTTTRAGTTT) (8). Resulting amplicons underwent purification and were quantified before being pooled for sequencing. High-throughput sequencing was then conducted on an Illumina HiSeq 2500 platform at Novogene in Guangzhou City, Guangdong Province, China.
To ensure the quality of downstream analyses, raw paired-end reads were filtered to eliminate those containing three or more ambiguous nucleotides. We also removed reads with a low average quality score (<20) and short reads (<100 nt), including those associated with barcodes, to generate clean concatenated reads that spanned the complete V4–V5 region of the 16S rRNA gene. High-quality sequences were subsequently trimmed and demultiplexed using the Quantitative Insights Into Microbial Ecology (QIIME1) pipeline (9). Operational taxonomic units (OTUs) were identified employing the unoise3 algorithm, which is a k-mer-based approach for sequence identification and error correction. OTUs derived from chloroplasts and mitochondria (approximately 1%), as well as singleton OTUs, were removed from the final OTU dataset (10). The Greengenes database was utilized for taxonomic classification with a confidence threshold of 90%.
-
Statistical analyses were executed utilizing SPSS 20 (SPSS Inc., Chicago, USA), with a significance threshold set at P<0.05 for all tests. Principal Coordinate Analysis (PCoA) was conducted to ascertain the global distribution patterns of ARGs and bacteria across the samples, applying the Bray-Curtis dissimilarity metric, facilitated by Canoco version 5.0. The significance of differences in microbial community compositions between treatment groups was assessed using Adonis, implemented in the R environment utilizing the “vegan” package (11). Additionally, heatmaps were generated in R employing the “pheatmap” package to visually represent the data (12). The relationships between ARGs and MGEs patterns were determined using ordinary least squares (OLS) regression models (13). Moreover, the Mantel test was applied for correlation analysis, leveraging the capabilities of R Studio in conjunction with the “vegan” package.
Microcosm Design and Sample Collection
DNA Extraction and HT-qPCR
Bacterial 16S rRNA Gene Sequencing and Bioinformatic Processing
Statistical Analysis
-
The enumeration of ARGs and MGEs varied across samples, with a range of 28 to 38 detected in each (Figure 2A). Notably, the quantity of ARGs present in plant roots and leaves was less than that found in the corresponding rhizosphere soil. Furthermore, when comparing treatments, it was observed that seed dressing and soil application of CeO2 NPs notably reduced the abundance of ARGs within the rhizosphere soil, in contrast to treatments using reclaimed water containing CeO2 NPs.
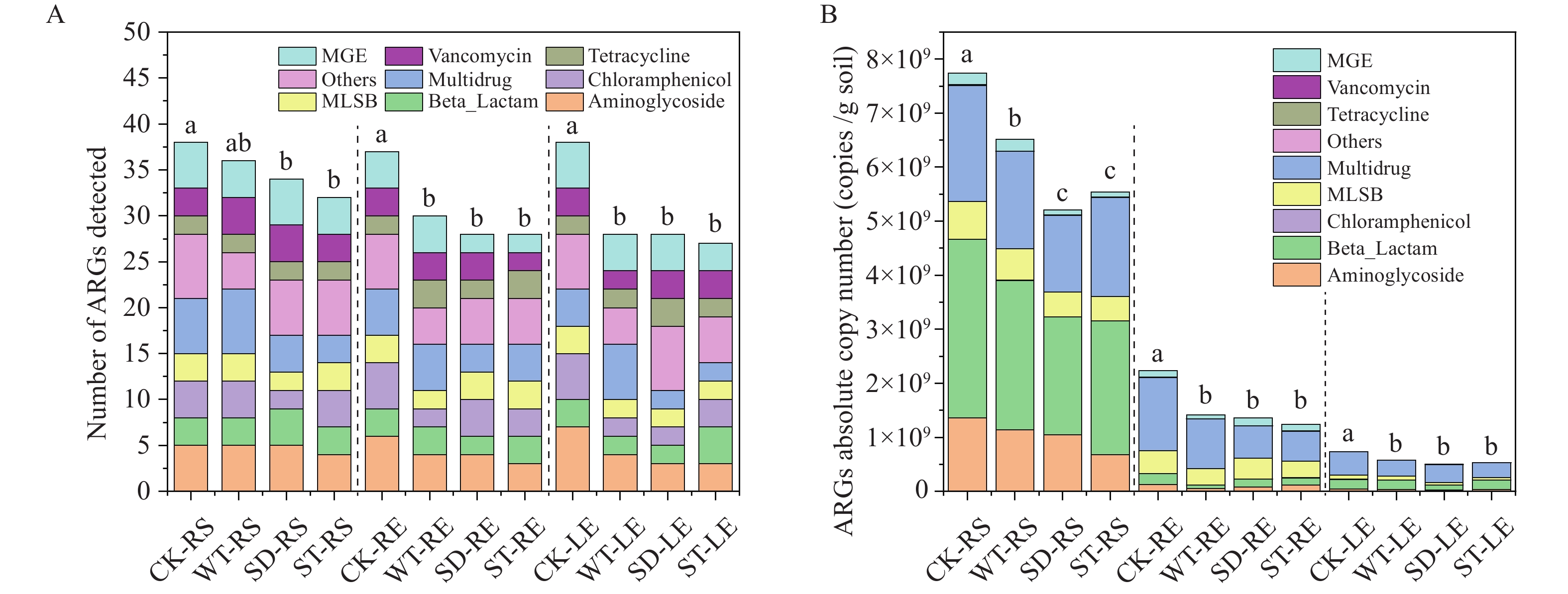 Figure 2.
Figure 2.The number and absolute abundances of ARGs genotypes. (A) Number of ARGs genotypes detected in different samples. (B) Absolute abundances of ARGs expressed as copies per g solid.
Abbreviation: ARGs=antibiotic resistance genes; MGE=mobile genetic element; MLSB= macrolide-lincosamide-streptogramin B.The concentration of ARGs in the soil, which ranged from 5.21×109 to 7.74×109 copies/g soil, was approximately an order of magnitude higher than that observed in the phyllosphere, with values spanning from 5.04×108 to 7.31×108 copies/g plant tissue (Figure 2B). Moreover, the concentration of ARGs in root endophytes (ranging from 1.25×109 to 2.36×109 copies/g plant) was significantly lower than the concentration found in rhizosphere soil.
To ensure that variations in the native bacterial populations did not confound our results, we normalized the abundance of ARGs to the number of bacterial cells, quantified as 16S rDNA copy numbers (
Supplementary Table S1 ). Given that the current average number of 16S rRNA genes per bacterial cell is approximately 4.1, the normalized ARG copy numbers varied from 0.10 to 0.35 copies per bacterial cell. The application of NPs had a discernible impact on the normalized abundance of ARGs, mirroring the effect seen with the absolute ARG copy numbers, with the exception of the rhizosphere soil subjected to the WT treatment (Supplementary Table S1 ). -
The abundance of mobile genetic elements (MGEs) exhibited considerable variability, spanning more than an order of magnitude, with counts ranging from 1.79×107 to 1.21×108 copies per gram of solid matter in the phyllosphere, and from 8.57×108 to 5.54×109 copies per gram of solid matter in the soil (Figure 2B). Similarly, reflecting the observed decline in the abundance of ARGs, the introduction of NPs was associated with a consequent reduction in the prevalence of MGEs within the water-soil-plant continuum.
-
If ARGs are identified solely in the roots or leaves of radishes when irrigated with reclaimed water, these ARGs are considered transmissible within the reclaimed water-soil-radish pathway. As depicted in Figure 3A, there are 30 ARGs capable of moving from reclaimed water to the leaves of radishes under the control condition. Relative to the control condition, we observed a reduction of 30.0%, 43.4%, and 40.0% in the number of transmissible ARGs from reclaimed water to radish leaves when NP applications in reclaimed water, seed dressing, and soil mixing interventions were implemented, respectively (Figure 3B–D).
 Figure 3.
Figure 3.Venn diagrams depicting the dissemination of ARGs from reclaimed water to radish seedlings. (A) control treatment, (B) water treatment, (C) seed dressing, and (D) soil treatment with NPs. (E) The fold change of the main 21 ARGs and MGEs which can spread from reclaimed water to radish.
Note: The outermost light blue circle represents the number of ARGs detected in reclaimed water, while the inner brown, green, and red circles represent the number of ARGs detected in soil, leaf endophytic, and root endophytic proportions, respectively. The dark blue circle in the middle represents the number of ARGs that can propagate in the reclaimed water-soil-radish system.
Abbreviation: NPs=nanoparticles; ARGs=antibiotic resistance genes; RW=reclaimed water; MGE=mobile genetic element; MLSB=macrolide-lincosamide-streptogramin B.
After the application of NPs, the majority of ARGs exhibited a decreasing trend in abundance. The aadA2 gene, associated with aminoglycoside resistance, exhibited the most substantial reduction, with a 48% decrease in the ST-RE group as shown in Figure 3E. Conversely, there was an upward trend observed in the abundance of vancomycin resistance genes, tetracycline resistance genes, and mexB, which is linked to multidrug resistance. Two ARGs, pbrT and qepA, showed a significant increase in the radish root when subjected to seed dressing and soil treatments. Following the NPs treatments, the abundance of three MGEs (Tn403, TrbC, and IS256) showed a significant decrease, particularly in the WT-RS, WT-LE, and SD-LE treatments.
-
Upon merging overlapped read pairs and conducting quality filtering, we obtained a total of 12,412 high-quality sequences from all the samples. These sequences were subsequently denoised, yielding 1,219 OTUs. We organized the microbial communities into 17 phyla by using the 16S rRNA database (GreenGene, gg_13_5 version). Principal coordinate analysis (PCoA) at the genus level indicated distinct separation of soil, root, and leaf sample communities along the PC1, PC2, and PC3 axes, which accounted for 30.5%, 22.6%, and 5.3% of the variation, respectively (Figure 4A). The PCoA further revealed pronounced clustering of bacterial communities that correlated with the application of different NPs in the root and leaf samples. This finding suggests that NP application significantly influences the overall composition of bacterial communities in both roots and leaves.
 Figure 4.
Figure 4.Principal coordinate analysis (PCoA) based on genus level communities profile (using Bray-Curtis distance). (A) overall, (B) rhizosphere soil, and (C) root and (D) leaf endophytic proportions of radishes.
Understanding the specific members of a community that drive bacterial population shifts is crucial. As indicated in
Supplementary Tables S2 –S4 , the genus Pseudomonas plays a pivotal role in the bacterial community dynamics of rhizosphere soil, as well as radish roots and leaves. Intriguingly, Pseudomonas was the taxonomic group that exhibited a significant decline across various NPs application methods in both soil and radish samples (Supplementary Figure S1 ). This reduction can likely be attributed to the pronounced antimicrobial properties of NPs and their ability to inhibit biofilm formation. It appears that NPs may reduce the competitive edge of certain bacteria, forcing them to relinquish their ecological niches and leading to an alteration in the structure of the bacterial community. -
The absolute abundance of MGEs was found to be linearly and positively correlated with the absolute abundance of ARGs ( R2=0.64, P<0.001) (Figure 5A). This indicates that the variation within the antibiotic resistome of reclaimed water-soil-plant systems is closely associated with MGEs. Figure 5B–D demonstrates that Procrustes analysis, utilizing Bray-Curtis dissimilarity metrics, confirmed a significant correlation between the abundance of ARGs and bacterial diversity within reclaimed water-soil-plant systems (P<0.01). Considering the observed community variations attributed to NP application (Figure 4), we postulate that NPs could potentially reduce the risk of ARG transmission in these systems by fostering a more beneficial community structure. As illustrated in Figure 6, microbial communities primarily facilitate the transmission of ARGs targeting beta-lactam, aminoglycoside, and tetracycline antibiotics, with considerable statistical significance (P<0.01, R>0.8). MGEs are chiefly associated with the dissemination of ARGs against aminoglycoside, multidrug, and macrolide-lincosamide-streptogramin B (MLSB) antibiotics. The regulation of aminoglycoside ARGs appears to be governed not only by bacterial communities but also by MGEs. Specifically, the aadA2 gene, associated with aminoglycoside resistance, exhibited the most pronounced decrease following silver nanoparticle treatment (Figure 3E). This suggests that the reduction may result from the combined influence of microbial community structure and horizontal gene transfer mechanisms. Conversely, the tetracycline resistance genes tetG and tetR increased under WT and SD conditions, leading us to conjecture that microbial communities play a vital role in this dynamic. Overall, the findings indicate that modifications in the resistome of reclaimed water-soil-plant systems are determined by the interplay of bacterial communities and horizontal gene transfer, working in concert to shape antibiotic resistance patterns.
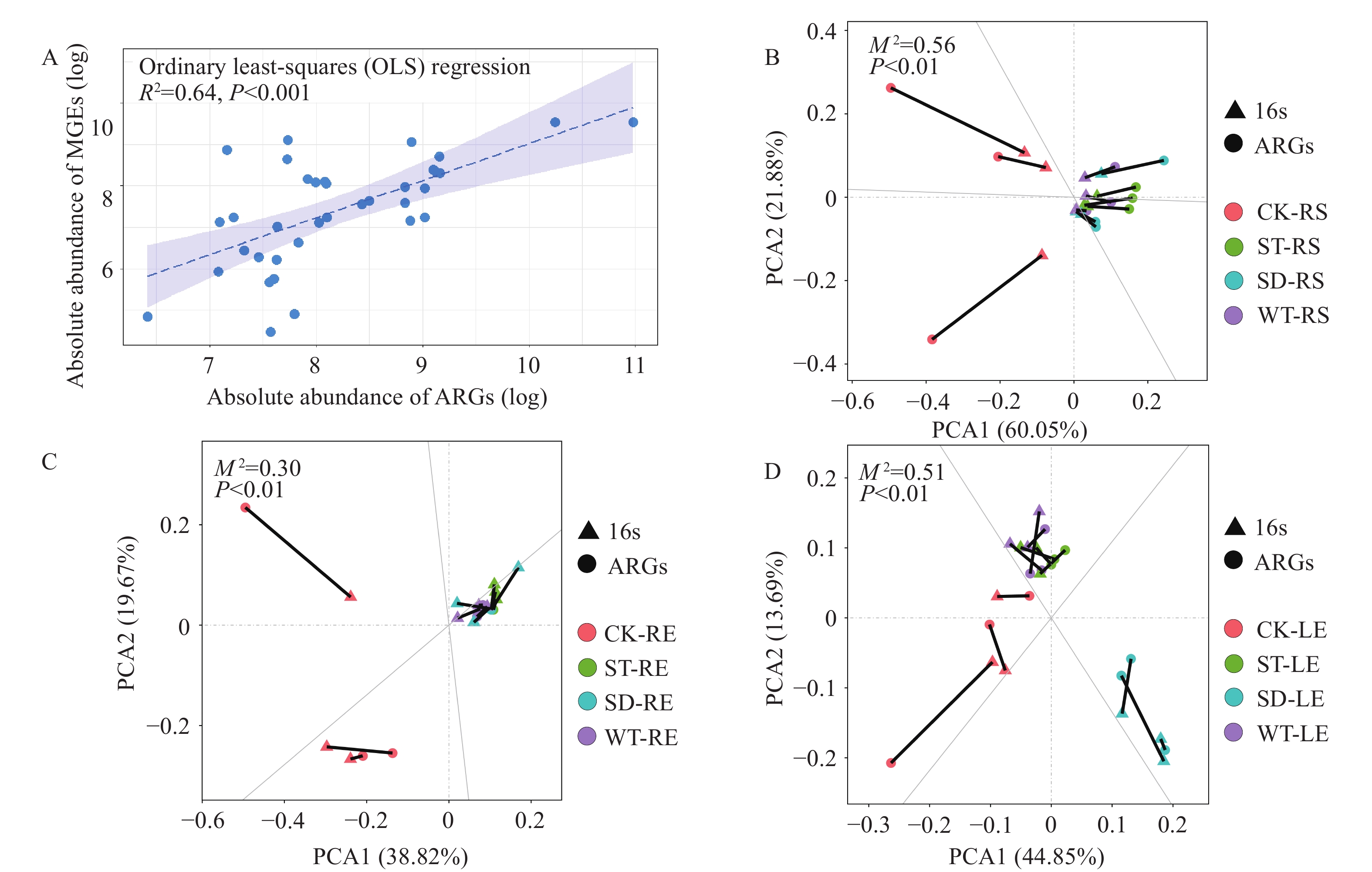 Figure 5.
Figure 5.Correlation analysis of ARGs with MGEs and bacterial community. (A) Ordinary least squares (OLS) regression showing the relationship between total absolute abundance of ARGs and MGEs. (B–D) Procrustes analysis depicts the correlation between ARG content and bacterial community in different samples (B, Rhizosphere soil; C, Root endophytic; D, Leaf endophytic).
Abbreviation: ARGs=antibiotic resistance genes; MGEs=mobile genetic elements; 16s=16S ribosomal RNA. Figure 6.
Figure 6.Pairwise comparisons of ARGs.
Note: 16s (OTUs) and MGEs were related to each ARG by partial (geographic distance–corrected) Mantel tests. Edge width corresponds to Mantel’s R statistic for the corresponding distance correlations, and edge color denotes the statistical significance.
Abbreviation: Beta=beta-lactams; Amin=aminoglycoside; Multi=multidrug; Van=vancomycin; Tet=tetracycline; MLSB=macrolide-lincosamide-streptogramin B; Chl=chloramphenicol.
Composition of ARGs in Reclaimed Water-Soil-Plant System
Composition of MGEs in reclaimed water-soil-plant system
Distribution of ARGs in Reclaimed Water-Soil-Plant System
Microbial Community Assembly
Bacterial diversity and MGEs accounting for antibiotic resistome
-
The utilization of NPs significantly impacted the prevalence of ARGs within reclaimed water (RW)-soil-plant systems, as demonstrated by a marked reduction in the absolute copy number of ARGs (P<0.05). This phenomenon may be explained by two potential mechanisms: (1) NP application modified the structure of the bacterial community, and (2) NP application suppressed the horizontal gene transfer of ARGs.
The resistome exhibited a significant correlation with the structures of bacterial communities (7). Our research findings indicate that different NP application methods can elicit distinct alterations in the microbial community structure of radishes, which suggests that these methods may influence the crop's resistome in varied ways. Intriguingly, our results also suggest that the modulation of bacterial communities by NPs tends to decrease the abundance of ARGs within plant microbiomes. This phenomenon may be attributable to the suppressive effects of NPs on biofilm formation, which serves as a nexus for ARG transmission. Furthermore, NPs may also impede the HGT of ARGs. HGT constitutes a primary pathway for the dissemination of ARGs in the environment, involving the exchange of genetic material among different bacterial species. Studies have shown that NPs can obstruct the transfer of plasmids — small, circular DNA molecules capable of carrying ARGs. This obstruction has the potential to thwart the inter-species propagation of ARGs, thereby curtailing their prevalence.
Alterations in the structure of endophytic bacterial communities may be linked to the varied levels of CeO2 NP uptake by plants, a process influenced by the differing NP application techniques (
Supplementary Figure S2 ). These results suggest that the method of NP administration can cause fluctuating NP absorption rates in plants. When compared to WT and SD treatments, the ST demonstrated a more pronounced inhibitory effect on the transmission of ARGs. This effect is likely due to the more efficient NP uptake by radishes in the ST treatment. Given that plants predominantly absorb nutrients via their roots from deeper soil layers, it can be inferred that the more homogeneously integrated CeO2 NPs in the ST-treated soil facilitated their uptake by the radishes. Previous studies have shown that different NP concentrations can induce varying levels of bacterial community modifications. Consistent with this, our prior research has established that varying NP concentrations lead to different impacts on ARG dissemination. Therefore, it is hypothesized that variations in NP uptake by radishes, which arise from the use of diverse NP application methods, primarily drive resistome changes within the radish. Our earlier work has demonstrated that NPs can enhance the development of the root exodermis barrier, hindering the absorption of exogenous bacteria via water transport pathways (14). Lignin content is commonly used as a marker for root lignification, indirectly indicating the maturity of the root's extracellular barrier (15). However, we detected no substantial changes in radish root lignin levels following CeO2 NP addition (Supplementary Figure S3 ). Although CeO2 NPs did not promote radish growth (Supplementary Figure S4 ), the literature highlights their significant potential in reducing vegetable wilt diseases. Consequently, the utilization of CeO2 NPs in reclaimed water irrigation emerges as a promising strategy for controlling diseases in vegetable crops.In conclusion, our research indicates that the utilization of NPs can significantly diminish the prevalence of ARGs in reclaimed water-soil-plant systems. The observed reduction in ARG abundance can be linked to the unique characteristics of NPs, which modify the composition of the bacterial community and suppress the horizontal transfer of ARGs. Our findings suggest that soil treatment with NPs (ST group) enhances plant uptake of these particles, resulting in changes to the microbial community and regulation of gene transfer. Although CeO2 NPs may not visibly affect the phenotypical parameters of radishes, they potentially provoke metabolic changes at the subcellular level, such as increased disease resistance, among other responses. This study underscores the promising role of nanotechnology in diminishing ARGs in agricultural settings. Continued research is warranted to refine nanoparticle applications for this purpose.
-
No conflicts of interest.
| Citation: |

|