

 Download:
Download:




-
The Omicron variants of severe acute respiratory syndrome virus (SARS-CoV-2), first identified in Botswana and South Africa in November 2021, rapidly became the predominant global strain, supplanting the Delta variant and presenting several sub-strains, including BA.2, BA.4, and BA.5 (1). These sub-strains exhibited numerous mutations and exhibited substantial immune evasion capabilities, increasing the potential for recurrent Omicron infections (2). Multiplex real-time reverse transcription-polymerase chain reaction (rRT-PCR) has been employed to detect SARS-CoV-2 variants with S-gene mutations (3-4). However, rRT-PCR assays can be time-consuming and necessitate specialized equipment and high-standard laboratories that many developing countries lack.
As a result, the clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated protein (CRISPR/Cas) system-based nucleic acid detection method has been adopted for practical application. The CRISPR/Cas12a assay relies on the protospacer adjacent motif (PAM) sites (5’-TTTN-3’), formed by mutations at base 9,866 on the ORF1ab gene of the BA.2 variant, base 27,788 on the ORF7b gene of the BA.4 variant, and base 26,529 on the M gene of the BA.5 variant. Additionally, specific single-base mutations occurred at base 23,040 on the S gene of the BA.2 variant and base 27,889 of the BA.5 variant, both of which contained a nearby PAM site. These mutations can be utilized for the precise detection of BA.2, BA.4, and BA.5 variants.
In this study, we designed and analytically validated a method combining reverse transcription-recombinase polymerase amplification (RT-RPA) and CRISPR/Cas12a technology to differentiate SARS-CoV-2 Omicron BA.2, BA.4, and BA.5 variants from one another.
-
In this study, the specificity of the RT-RPA-CRISPR/Cas12a assay was assessed using wild-type SARS-CoV-2, its variants of concern (VOCs) including Delta and Omicron (BA.1, BA.2, BA.4, and BA.5), as well as influenza strains H7N9, PR8, and X31. Viral nucleic acids were prepared in the laboratory for the assay. Beijing CDC provided twenty samples each of BA.2 and BA.5, while Shenzhen CDC supplied six BA.4 samples. Additionally, negative clinical samples were obtained from staff at China CDC.
-
Specific mutation sites for the Omicron BA.2, BA.4, and BA.5 variants were identified using the covSPECTRUM webpage and mutation site information provided by the World Health Organization (WHO) (5-7). The reference genome sequence for SARS-CoV-2 was acquired from the National Centre for Biotechnology Information (NCBI) under the accession number NC_045512.2, and information on mutations in SARS-CoV-2 variants was sourced from Global Initiative on Sharing All Influenza Data (GISAID) (https://www.gisaid.org/). Genomic sequences of BA.2, BA.4, and BA.5 with GISAID accession numbers 12386761, 13651679, and 14253379, respectively, were used as reference sequences for designing RT-RPA primers and crRNA. The BA.2-specific nucleotide mutation sites (C9866T and A23040G), the BA.4-specific nucleotide mutation site (G27788T), and the BA.5-specific nucleotide mutation sites (G26529A and C27889T) were selected for further identification of crRNA recognition sequences containing a PAM sequence with 18 or 20 nucleotides. The resulting RT-RPA amplification products ranged from 100–500 base pairs in length (Figure 1).
 Figure 1.
Figure 1.Sequence alignment of the SARS-CoV-2 reverse transcription-recombinase polymerase-amplified target region genes with BA.2, BA.4, and BA.5 variants.
Note: The reference sequence used is NC_045512.2 SARS-CoV-2 strain. This figure displays the nucleotide positions of the reference genome amplification target region within the whole-genome sequence. Dots represent nucleotides identical to the reference genome sequence, with mismatched sequences denoted by "a", "t", "c," or "g". Arrows indicate the direction and sequence of upstream primers, downstream primers, and CRISPR-RNA (crRNA) recognition sites. -
Reverse transcription isothermal amplification was performed using a commercial RT-RPA kit (AMP-Future Biotech Co. Ltd., Weifang, China). There was 29.4 μL of Buffer A, 2 μL of forward and reverse primers (10 μmol/L) (Table 1), 5 μL of the sample, 2.5 μL of Buffer B, and 9.1 μL of nuclease-free in a 50 μL reaction mixture. Thermal cycling was performed at 42 °C for 30 min.
Gene/nucleotide mutation site Primers/crRNA and template sequence Sequence (5’–3’) C9866T RT-RPA-F CTAAAGTTGCGTAGTGATGTGCTATTACCT RT-RPA-R TCAGAACCTGAGTTACTGAAGTCATTGAGA crRNA-9866 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCGCAAUAUAAUAGAUACU crRNA-9866 template AGTATCTATTATATTGCGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA A23040G RT-RPA-F TACCTGTATAGATTGTTTAGGAAGTCTAAT RT-RPA-R AAAAGAAAGTACTACTACTCTGTATGGTTG crRNA-23040 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCGAACAUAUGGUUUCCGA crRNA-23040 template TCGGAAACCATATGTTCGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-23040-1 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCGAUCAUAUGGUUUCCGA crRNA-23040-1 template TCGGAAACCATATGATCGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-23040-2 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCGAACAUAUGGUUUCCGACC crRNA-23040-2 template GGTCGGAAACCATATGTTCGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-23040-3 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCGAUCAUAUGGUUUCCGACC crRNA-23040-3 template GGTCGGAAACCATATGATCGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA G27788T RT-RPA-F TTGCGGCAATAGTGTTTATAACACTTTGCTTC RT-RPA-R ATTTCATGTTCGTTTAGGCGTGACAAGTTTCA crRNA-27788 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCUUUUUAGCCUUUCUGUU crRNA-27788 template AACAGAAAGGCTAAAAAGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA G26529A RT-RPA-F TAGAGTTCCTGATCTTCTGGTCTAAACGAA RT-RPA-R GAAGACAAATCCATGTAAGGAATAGGAAAC crRNA-26529 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUCCAUGGCUAAAAUUAAAGUU crRNA-26529 template AACTTTAATTTTAGCCATGGATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-26529-1 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUGGCAUGGCAAAUUUCCAAC crRNA-26529-1 template GTTGGAATTTGCCATGCCATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-26529-2 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUGCCUUGGCAAAUUCCAAC crRNA-26529-2 template GTTGGAATTTGCCAAGGCATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA crRNA-26529-3 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUGGCAUGGCAAAUUCCAACGG crRNA-26529-3 template CCGTTGGAATTTGCCATGCCATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA C27889T RT-RPA-F TTATGCTTATTATCTTTTGGTTCTCACTTG RT-RPA-R AGGTGCTGATTTTCTAGCTCCTACTCTAAT crRNA-27889 UAAUACGACUCACUAUAGGGUAAUUUCUACUAAGUGUAGAUAAGUUCAUUUAGGCGUGACA crRNA-27889 template TGTCACGCCTAAATGAACTTATCTACACTTAGTAGAAATTACCCTATAGTGAGTCGTATTA Table 1. List of the reverse transcription-recombinase polymerase amplification (RT-RPA) primers and CRISPR-RNA (crRNA) sequences.
-
The crRNA sequences were designed for the CRISPR/Cas12a assay (Table 1) (8). crRNAs were transcribed by annealing DNA oligonucleotides (T7-gRNA-oligonucleotide), which contained a T7 promoter, conserved stem-loop sequences, and guide sequences with a specific DNA oligonucleotide. The synthesis of crRNAs was conducted at 37 °C for 4 hours using a RiboMAX™ Large Scale RNA Production System-T7 Kit (Promega, Madison, WI, USA). The resulting crRNA was then purified utilizing the RNeasy Mini Kit (Qiagen, Hilden, Germany) and assessed for purity and concentration using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Finally, samples were aliquoted and stored at −80 °C.
-
Cas12a-mediated target cleavage assays were conducted in a 22 μL reaction volume, using 50 nmol/L LbCas12a (Tolo Biotech, Shanghai, China) preincubated with 100 nmol/L crRNA in 1×TOLOBIO buffer at 37 °C for 10 minutes to form a crRNA-Cas12a complex. Subsequently, 5 μL of RT-RPA product and 500 nmol/L probe reporter (5’-6-FAM-TTATT-BHQ-1-3’) were added, and the mixture was incubated at 37 °C for 20 minutes. The fluorescence signal was monitored every 30 seconds using a fluorescent detector (Kunpeng, Beijing, China). Additionally, the fluorescence signal was scanned with an Amersham ImageQuant 800 (Cytiva, Marlborough, MA, USA).
-
Nucleic acid quantification for SARS-CoV-2 was conducted utilizing a digital PCR instrument (Sniper Medical Technology Ltd., Jiangsu, China), with an assay developed in our laboratory (9). Samples were examined at four dilutions (ranging from 1,000 to 1 copies/μL) in order to determine concentration levels based on the N gene.
-
The RT-RPA-CRISPR/Cas12a assay yielded positive results when fluorescence levels between cycle 1 and cycle 40 were at least 1.5 times greater than those of the negative control; otherwise, the assay was considered negative. Data processing and graphical representation were performed using GraphPad Prism software (GraphPad, Boston, MA, USA).
-
We developed crRNA-9866 to identify the BA.2 variant (Figure 2A). Since Cas12a detection relies on the presence of a PAM site, the targeted locus 9,866 features a single-nucleotide C/T mutation at position nt9866 in the SARS-CoV-2 ORF1ab gene. This leads to the emergence of the PAM sequence in the BA.2 variant, which subsequently activates the trans-cleavage activity of Cas proteins and cleaves fluorescent reporter molecules. We employed this characteristic to differentiate BA.2 strains; however, this site exhibited cleavage activity for both BA.2 and other SARS-CoV-2 strains, without demonstrating specificity (Figure 2A).  Figure 2.
Figure 2.Specific identification of SARS-CoV-2 BA.2, BA.4, and BA.5 strains using Cas12a cis-cleavage and trans-cleavage by specific CRISPR-RNA (crRNA). (A) In vitro cleavage assay of C9866T targets using an 18 bp crRNA-9866 to detect BA.2 strains. (B) In vitro cleavage assay of A23040G targets using crRNA-23040 to detect BA.2 strains. An 18 bp crRNA-23040 by introducing a “T to A” transition mismatched at position 4 of the crRNA. (C) In vitro cleavage assay of A23040G targets using an 18 bp crRNA-23040-1 to detect BA.2 strains. (D) In vitro cleavage assay of A23040G targets using crRNA-23040-2 to detect BA.2 strains. A 20 bp crRNA-23040-2 by introducing a “T to A” transition mismatched at position 4 of the crRNA. (E) In vitro cleavage assay of A23040G targets using a 20bp crRNA-23040-1 to detect BA.2 strains. (F) In vitro cleavage assay of C27788T targets using an 18 bp crRNA-27788 to detect BA.4 strains. (G) In vitro cleavage assay of G26529A targets using a 20bp crRNA-26529 to detect BA.5 strains. (H) In vitro cleavage assay of G26529A targets using crRNA-26529-1 to detect BA.5 strains. An 18bp crRNA-26529-1 by introducing a “C to G” transition mismatched at position 2 of the crRNA. (I) In vitro cleavage assay of G26529A targets using crRNA-26529-2 to detect BA.5 strains. An 18 bp crRNA-26529-2 by introducing an “A to T” transition mismatched at position 4 of the crRNA. (J) In vitro cleavage assay of G26529A targets using crRNA-26529-3 to detect BA.5 strains. A 20 bp crRNA-26529-3 by introducing a “C to G” transition mismatched at position 2 of the crRNA. (K) In vitro cleavage assay of C27889T targets using crRNA-27889 to detect BA.5 strains. A 20 bp crRNA-27889 by introducing a “T to A” transition mismatched at position 2 of the crRNA. (L) Verification of the specificity of crRNA-23040-2 for BA.2 with other variants of SARS-CoV-2 and influenza viruses. (M) Verification of the specificity of crRNA-27788 for BA.4 with other variants of SARS-CoV-2 and influenza viruses. (N) Verification of the specificity of crRNA-26529-2-27889 with other variants of SARS-CoV-2 and influenza viruses.
Abbreviation: WT=wild-type nucleic acids; AU=Arbitrary Unit; NTC=negative control.We designed an 18 bp-long crRNA by introducing a transition mismatch in crRNA-23040 at position 4, which yielded negative results (Figure 2B). To enhance efficiency, we designed an unmodified transition mismatch crRNA-23040-1 at position 4, along with a 20 bp-long crRNA line consisting of both a transition mismatch in crRNA-23040-2 and an unmodified transition mismatch in crRNA-23040-3 at the same position. The results demonstrated that crRNA-23040-1 and crRNA-23040-3 detected both BA.2 and other subtypes of SARS-CoV-2 strains, thus displaying no specific cleavage ability (Figure 2C, E). However, BA.2 exhibited significant cleavage in the presence of crRNA-23040-2, while no cleavage was observed among other SARS-CoV-2 subtypes (Figure 2D, L). These findings indicate that crRNA-23040-2 can specifically identify BA.2 variants.
The locus 27,788 is distinguished by a single-nucleotide mutation (G/T) at position nt27788 in the SARS-CoV-2 ORF7b gene, which generates the PAM sequence in the BA.4 lineage. Consequently, the crRNA-27788 was designed to specifically detect the BA.4 lineage from other SARS-CoV-2 strains. Significant cleavage was observed for BA.4 in the presence of crRNA-27788, while no cleavage was detected for other SARS-CoV-2 subtypes (Figure 2F, M). These results suggest that crRNA-27788 can accurately and specifically identify BA.4 variants.
The BA.5 variant exhibits a mutation at nt26529, resulting in a PAM locus (Figure 2G). To design an 18 bp length, carefully mismatched crRNA-26529-1 and 26529-2 were introduced at positions 2 and 4, respectively, while a 20 bp length was introduced at position 2 for mismatched crRNA-26529-3 and crRNA-27889 (Figure 2H–K). Subsequent analyses revealed that crRNA-26529-2 and crRNA-27889 specifically recognized the BA.5 variant; thus, they were chosen for further evaluation (Figure 2I, K). The CRISPR-Cas12a mediated triple-line lateral flow assay, coupled with multiplex RT-RPA reactions, has been suggested as a platform for rapid, simultaneous two-gene detection of SARS-CoV-2 in point-of-care testing, which could significantly enhance detection accuracy and efficiency (10). Consequently, to improve detection efficiency, crRNA-26529-2 and crRNA-27889 were combined to form crRNA-26529-2-27889 for dual-gene detection of BA.5 (Figure 2N). This demonstrates that crRNA-26529-2-27889 can specifically identify BA.5 variants.
-
The crRNA-23040-2, crRNA-27788, and crRNA-26529-2-27889 were chosen for the final evaluation of the assay due to their enhanced sensitivity. The respective limits of detection for these crRNAs were 10, 1, and 10 copies/μL (Figure 3). Fluorescence readings demonstrated consistency across the three replicate experiments, and the limits of detection exhibited strong reproducibility (Figure 3G–I).
 Figure 3.
Figure 3.Sensitivity and repeatability evaluation. (A–C) LOD for Cas12a fluorometric detection method. (D–F) LOD for Cas12a imager-based detection method. (G–I) Results from three replicate experiments, showing mean fluorescence values obtained in three independent trials.
Abbreviation: AU=Arbitrary Unit; cp=copies; NTC=negative control; LOD=limit of detection. -
A total of 46 SARS-CoV-2 variant RNA samples, previously analyzed by next-generation sequencing, were examined using the established assay. Each sample was subjected to triplicate RT-RPA reactions in order to detect the presence of BA.2, BA.4, or BA.5 variants. The results demonstrated that the BA.4 and BA.5 assay systems accurately identified all BA.4 (n=6) and BA.5 (n=20) clinical samples with 100% sensitivity and specificity. In contrast, the BA.2 assay missed two out of 20 samples, yielding a sensitivity of 90% and specificity of 100% (Figure 4).
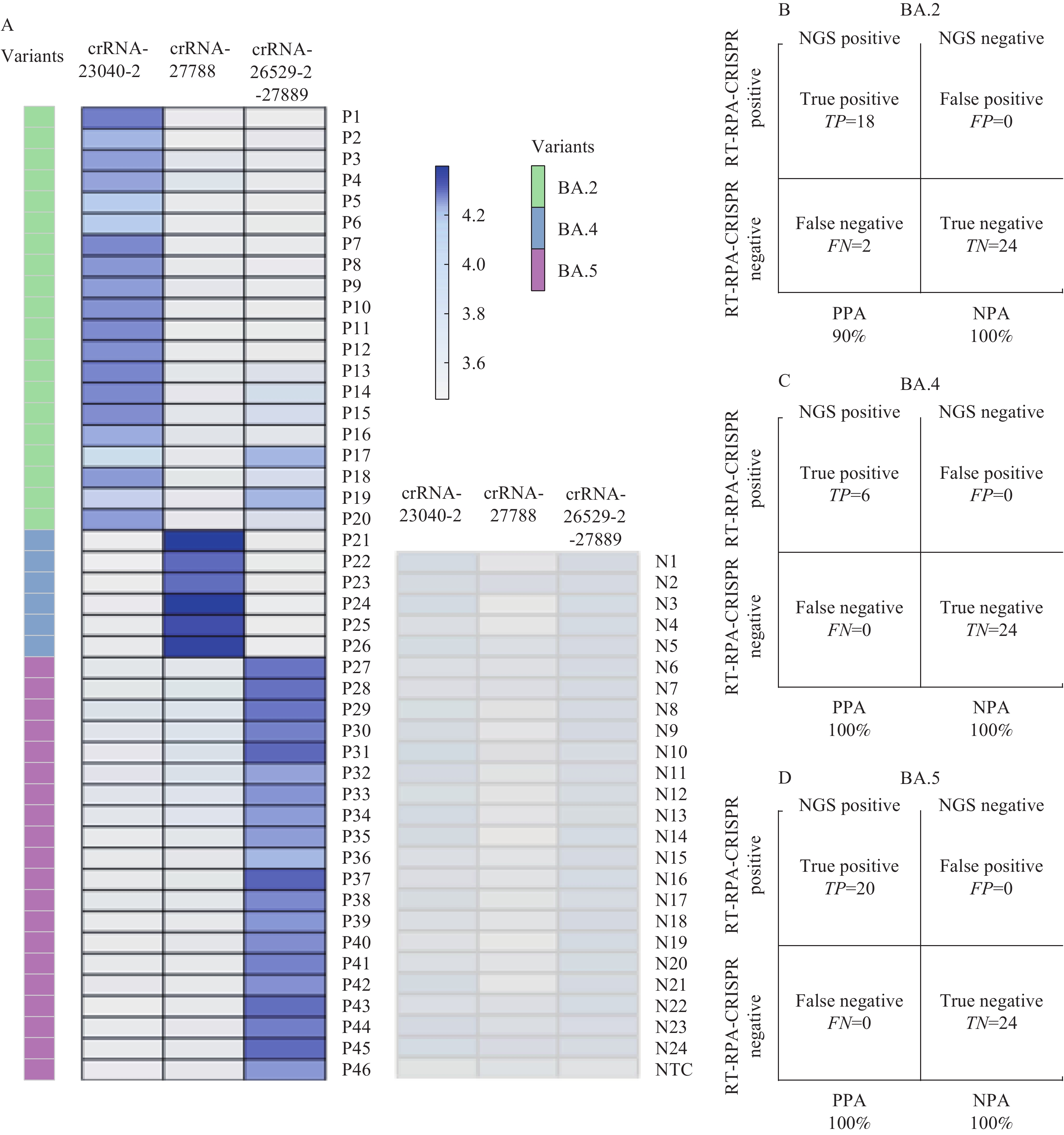 Figure 4.
Figure 4.Detection of SARS-CoV-2 BA.2, BA.4, and BA.5 in clinical samples using the CRISPR/Cas12a system. (A) Endpoint fluorescence readouts for the reverse transcription-recombinase polymerase amplification (RT-RPA-CRISPR) detection assay were obtained from clinical samples. (Left) Nasopharyngeal swab extracts from 46 positive and (Right) 24 negative patients, as determined by next-generation sequencing, were tested. Blue colors represent a positive result. (B) Sensitivity and specificity of the RT-RPA-CRISPR assay for detecting BA.2. (C) Sensitivity and specificity of the RT-RPA-CRISPR assay for detecting BA.4. (D) Sensitivity and specificity of the RT-RPA-CRISPR assay for detecting BA.5.
-
The sensitivity of crRNA detection for single-base mutations varies depending on the position and length of the mutations (11). Huang et al. demonstrated that the specificity of crRNA binding in the SARS-CoV-2 typing assay was closely correlated with the seed region of the crRNA when single-base mismatches were introduced (12). In the current study, we compared two BA.2 crRNAs of differing lengths (18 and 20 bp) with and without single-base mismatches. Results showed that crRNAs of both lengths, without mismatches, cleaved both BA.2 and other mutant strains, while 20 bp crRNAs with single-base mismatches specifically detected BA.2.
CRISPR/Cas detection depends on PAM sites, which can be mutated by the addition of PAM sites. In this study, the locus 27,788 mutation added to the PAM sites, facilitating BA.4 detection. For the BA.5 variant, two loci (26,529 and 27,889) were selected. Locus 27,889 was designed with a single-base mismatch in the 20 bp crRNA, while locus 26,529 was designed to introduce or omit a single-base mismatch at 18 bp and 20 bp, respectively. Both crRNA-26529-2 and crRNA-27889 specifically detected the BA.5 variant.
However, a preliminary assessment demonstrated that the BA.5 detection by crRNAs resulted in weak fluorescence intensity. Consequently, a dual-site detection approach was proposed, combining crRNA-26529-2 and crRNA-27889 as crRNA-26529-2-27889. The crRNA-26529-2-27889 detection of BA.5 variants exhibited high specificity and sensitivity.
The application of Cas12a-based techniques for detecting specific mutations in SARS-CoV-2 has been documented in recent literature. Liang et al.’s CRISPR-based genotyping approach can be employed for identifying the most VOC or variants of interest in SARS-CoV-2 (8,13). Fasching et al. targeted L452R, E484K/Q/A, and N501Y mutant loci within the S gene to identify the most widespread VOC or variant of interest, including those containing Omicron (14). However, only a limited number of methods or algorithms currently exist for genotyping SARS-CoV-2 Omicron subtype strains. The present study has revealed that CRISPR-based genotyping techniques are effective in classifying BA.2, BA.4, and BA.5 subtypes of SARS-CoV-2 Omicron strains.
The CRISPR system employed in this study effectively discriminated between mutations present in the SARS-CoV-2 BA.2, BA.4, and BA.5 variants, exhibiting high sensitivity. Additionally, a consistent level of sensitivity was observed through both fluorescence signal readout and imaging for the BA.2, BA.4, and BA.5 variants, surpassing that of the NASBA-based combined CRISPR-Cas13a locus detection system (N501Y; 82 copies/response) and the LAMP-CRISPR12 combined detection systems (L452R, E484K/Q/A, and N501Y; 10,000 copies/response) (14-15). Sequence analysis revealed that emerging BA.5.2.48, BF.7.14, and BQ.1.1 variants, all subvariants of BA.5, contain the detection sites targeted in our BA.5 analysis, indicating a theoretical potential for detection.
A key limitation of this study, however, was the absence of validation using a substantial set of clinical samples. Additionally, false positives were observed at site 27,889 in some BA.2 samples due to the crRNA seed sequence being specifically identified at alternate locations within the SARS-CoV-2 sequence when single-base mismatches were introduced. Further refinement of the method could enhance assay specificity.
-
No conflicts of interest.
HTML
Samples
Identifying RT-RPA Primers and CRISPR-RNA (crRNA) Recognition Sequences
RT-RPA Amplification
Preparation of crRNA
Cas12a-Mediated Assay
Digital PCR
Statistical Analysis and Interpretation of Results
Specificity Assessment
Sensitivity and Repeatability Evaluation
Evaluation of Cas12a Detection Assay Performance using Clinical Samples
| Citation: |

|