
 Download:
Download:




-
Foodborne antibiotic-resistant enterococci represent a critical surveillance priority within the “Human-Animal-Environment” One Health framework (1). Enterococcus faecium (E. faecium) exhibits a dual nature: it serves as a beneficial probiotic in fermented foods and dietary supplements while simultaneously acting as an opportunistic pathogen capable of causing antibiotic-resistant infections, including endocarditis and sepsis (2). In contrast, Enterococcus lactis (E. lactis) strains are generally regarded as non-pathogenic probiotic bacteria (3). Historically, phylogenetic analyses classified E. faecium into two distinct clades: Clade A, comprising clinical and animal isolates, and Clade B, consisting of strains from healthy humans (4). However, recent genomic studies demonstrated that Clade B isolates share >97% average nucleotide identity (ANI) with E. lactis, exceeding the 95% species delineation threshold and prompting their reclassification as E. lactis (5). Despite this taxonomic revision, comprehensive long-term surveillance data characterizing the antibiotic resistance (ABR) profiles and genomic features of redefined E. faecium and E. lactis populations within the Chinese food chain remain limited.
In this study, we collected samples from multiple nodes along the food chain across five provincial-level administrative divisions (PLADs) in China during 2015–2019 and 2023–2024. We conducted a comparative analysis of ABR phenotypes and genomic content between E. faecium and E. lactis isolates. This investigation aimed to elucidate the distinct ABR risks posed by these two species within the Chinese food chain and provide evidence-based data to support risk assessment and control strategies for foodborne antibiotic-resistant enterococci.
-
During 2015–2019, 694 samples were collected from Beijing, Henan, Hubei, and Jilin PLADs. From 2023–2024, 1,539 samples were collected from Beijing, Henan, Hubei, Jilin, and Hebei PLADs (
Supplementary Table S1 ). Samples were obtained from multiple sites and sources across the food chain, including food animals, food products, environmental surfaces, and human specimens, to capture the diversity of enterococcal populations. Sample collection, Enterococcus detection, and identification procedures followed previously established methods (1). -
Whole-genome sequencing of Enterococcus isolates was performed using the Illumina HiSeq platform (Illumina Inc., United States). Paired-end libraries (2×150 bp) were prepared with the Nextera DNA Sample Preparation Kit (Illumina Inc., United States) according to the manufacturer’s protocol. High-quality paired-end reads were processed using Shovill v1.1.0 (https://github.com/tseemann/shovill), and de novo genome assemblies were generated with SPAdes (version 3.15.5; Algorithmic Biology Lab, St. Petersburg, Russia), yielding contigs (6). ANI values were calculated for all assembled Enterococcus genomes using FastANI (version 1.3.3; Atlanta, USA), with a species delineation threshold of 95%(7).
-
Antimicrobial susceptibility of E. faecium and E. lactis isolates was determined by broth microdilution and interpreted according to Clinical & Laboratory Standards Institute (CLSI) interpretive criteria. Minimum inhibitory concentrations (MICs) were determined for 13 antibacterial compounds: ampicillin and penicillin (β-lactams), erythromycin (a macrolide), enrofloxacin and ciprofloxacin (quinolones), daptomycin and vancomycin (glycopeptides), tetracycline and doxycycline (tetracyclines), chloramphenicol (a phenicol), high-level gentamicin (HLGA) and high-level streptomycin (HLSA) (aminoglycosides), and linezolid (an oxazolidinone). An isolate was classified as multidrug-resistant (MDR) if it exhibited resistance to three or more antibacterial compounds from different classes.
-
Antibacterial resistance genes (ARGs), virulence factor encoding genes (VFs), mobile genetic elements (MGEs), and plasmid replicons were identified using ABRicate (https://github.com/tseemann/abricate) against the following databases: the Comprehensive Antimicrobial Resistance Database for ARGs (8), the Virulence Factors Database for VFs (9), the Mobile Orthologous Groups Database for MGEs (10), and PlasmidFinder (11) for plasmid replicons. Detection thresholds of 80% minimum coverage and 80% minimum identity were uniformly applied across all analyses.
To construct the core genome single nucleotide polymorphism (SNP)-based phylogenetic tree, the following bioinformatic pipeline was employed: Snippy v4.6.0 (https://github.com/tseemann/snippy) was used for core genome alignment and variant calling. Core genome alignment was reconstructed using Gubbins (version 2.4.1; Hinxton, UK) (12). A maximum likelihood phylogenetic tree was then inferred from non-repetitive core SNPs using FastTree (version 2.1.11; Berkeley, USA) with the GTR+CAT substitution model (13).
Roary (version 3.6.1; Exeter, UK) (14) was used to identify core genes and pan-genes. Pan-genome-wide association study (Pan-GWAS) was performed using Scoary (version 1.6.16; Oslo, Norway) (15). Species-specific genes were defined as those present in more than 80% of isolates from one species but absent in the other species. Clusters of Orthologous Groups (COG) classification of significantly associated genes was performed by mapping to the COG function database using BLAST.
-
Statistical analyses were conducted using SPSS Statistics for Windows, version 17.0 (SPSS Inc., Chicago, IL, USA). Categorical variables were compared using Pearson’s chi-squared test and Fisher’s exact test. Correlation analyses between dichotomous categorical variables were performed using the Phi coefficient. Statistical significance was defined as P<0.05.
-
As shown in
Supplementary Table S2 , E. lactis exhibited a significantly higher prevalence rate (6.9%, 153/2,233) in the food chain compared to E. faecium (3.9%, 87/2,233) (P<0.001). Additionally, E. lactis demonstrated significantly higher prevalence rates than E. faecium in both food and livestock samples (P<0.05). This pattern was particularly pronounced during 2023–2024, when E. lactis prevalence significantly exceeded that of E. faecium (P<0.001). Temporal analysis revealed that neither species exhibited significant changes in prevalence rates when comparing the 2015–2019 period to the 2023–2024 period (P>0.05). -
E. faecium isolates demonstrated significantly higher resistance rates to 12 antimicrobials compared with E. lactis (P<0.05) (Figure 1). In contrast, E. lactis exhibited a significantly higher resistance rate to erythromycin than E. faecium (P<0.01). Specifically, E. faecium showed elevated resistance to erythromycin (58/87, 66.7%), tetracycline (51/87, 58.6%), and enrofloxacin (48/87, 55.2%), while displaying minimal resistance to daptomycin (3/87, 3.4%), vancomycin (6/87, 6.9%), and linezolid (6/87, 6.9%). E. lactis exhibited the highest resistance rates to erythromycin (127/153, 83.0%), tetracycline (29/153, 19.0%), and ciprofloxacin (18/153, 11.8%), but demonstrated negligible resistance to vancomycin (0%), daptomycin (1/153, 0.7%), and ampicillin (1/153, 0.7%). The MDR rate of E. faecium (43/87, 49.4%) significantly exceeded that of E. lactis (16/153, 10.5%) (P<0.001). Among MDR E. faecium isolates, the largest proportion originated from hospital patients (19/43, 44.2%), whereas among MDR E. lactis isolates, the largest proportion was food-derived (7/16, 43.8%). The most common resistance profile in E. faecium was ciprofloxacin-enrofloxacin co-resistance (5/87, 5.7%), while E. lactis most frequently exhibited tetracycline-erythromycin co-resistance (10/153, 6.5%).
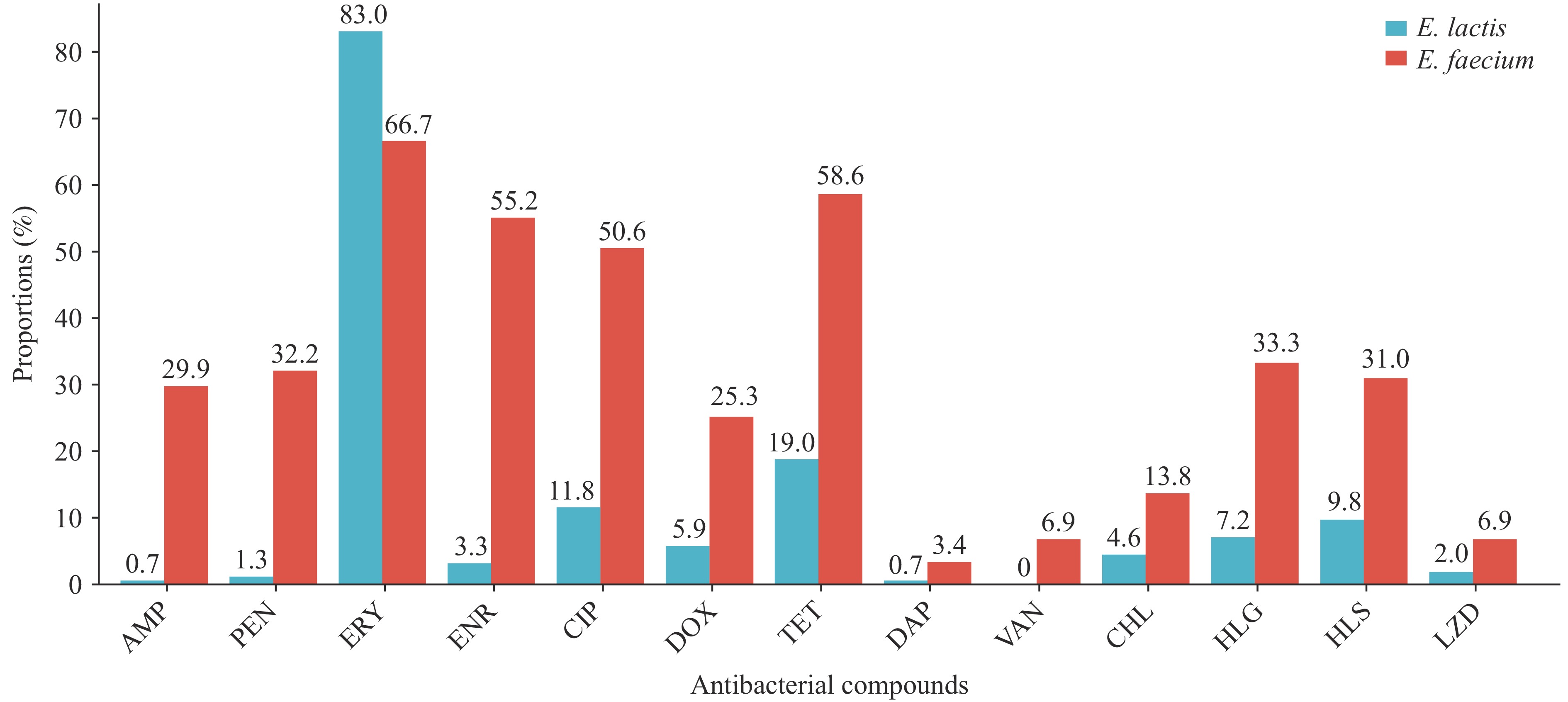 Figure 1.
Figure 1.Enterococcal resistance profiles against a panel of 13 antibacterial compounds.
Abbreviation: AMP=ampicillin; PEN=penicillin; ERY=erythromycin; ENR=enrofloxacin; CIP=ciprofloxacin; DOX=doxycycline; TET=tetracycline; DAP=daptomycin; VAN=vancomycin; CHL=chloramphenicol; HLGA=high-level gentamicin; HLSA=high-level streptomycin; LZD=linezolid; E. faecium=Enterococcus faecium; E. lactis=Enterococcus lactis.A total of 20 distinct ARG types belonging to 8 functional categories were detected across E. faecium and E. lactis genomes (Figure 2A). E. faecium carried significantly more ARGs than E. lactis (P<0.001). Furthermore, the tetM, tetL, ermB, and fexA genes in E. faecium showed significant associations with resistance phenotypes to their corresponding antimicrobials (tetracycline, erythromycin, and chloramphenicol) (Figure 2B). Notably, the macrolide resistance gene ermT in E. faecium demonstrated a significant association with resistance phenotypes to the two β-lactam antibacterials (ampicillin and penicillin). In E. lactis, the tetracycline resistance genes (tetL and tetM) exhibited significant correlations with resistance to both tetracycline antibacterials tested (doxycycline and tetracycline) (Figure 2C).
 Figure 2.
Figure 2.Phylogenetic relationships of E. faecium and E. lactis and correlation analysis between ARGs and ABR phenotypes. (A) Midpoint-rooted maximum likelihood phylogenetic tree constructed from 87 E. faecium genomes and 153 E. lactis genomes. (B) Correlation heatmap between ARGs and ABR phenotypes in E. faecium. (C) Correlation heatmap between ARGs and ABR phenotypes in E. lactis.
Note: For (A), reference strains used were SRR24 (accession number: GCA_009734005.2) for E. faecium and CX262 (accession number: GCA_019343125.1) for E. lactis. Colored bands on the right display genomic information for Enterococcus isolates. The first heatmap presents antibacterial susceptibility testing results for Enterococcus. The second heatmap illustrates the distribution of antibacterial resistance genes (ARGs) across Enterococcus isolates. The third heatmap depicts the distribution of virulence factor encoding genes in Enterococcus.
Abbreviation: ABR=antibacterial resistance; AMP=ampicillin; PEN=penicillin; ERY=erythromycin; ENR=enrofloxacin; CIP=ciprofloxacin; DOX=doxycycline; TET=tetracycline; DAP=daptomycin; VAN=vancomycin; CHL=chloramphenicol; HLGA=high-level gentamicin; HLSA=high-level streptomycin; LZD=linezolid; S=susceptible; I=intermediate; R=resistant; AST=antibacterial susceptibility testing; ARG=antibacterial resistance gene; E. faecium=Enterococcus faecium; E. lactis=Enterococcus lactis.
-
Core genome-based SNP phylogenetic analysis demonstrated distinct separation between E. faecium and E. lactis (Figure 2A). The two species carried comparable numbers of virulence factor genes, with no significant difference observed (P>0.05). We identified 25 unique VFs across four functional categories — adhesion, immune modulation, biofilm formation, and exotoxin production — in both E. faecium and E. lactis genomes. Adhesion genes predominated (18/26, 69.2%), followed by immune modulation factors (galE, cpsA, cpsB, and cpsD), biofilm formation genes (bopD and papR), and a single exotoxin gene (nheB). Three VFs — bopD (biofilm formation), cpsA, and cpsB (immune modulation) — were universally present across all isolates. The most prevalent adhesion genes were efaA (236/240, 98.3%), gapA (234/240, 97.5%), and acm (223/240, 92.9%). Notably, the adhesion gene sgrA exhibited differential distribution, appearing in 67.7% (20/30) of E. faecium isolates from hospital patients versus 83.0% (127/153) of E. lactis isolates. Comparative genomic analysis revealed that E. faecium harbored significantly higher numbers of ARGs, MGEs, and plasmid replicons compared to E. lactis (P<0.05) (Figure 3).
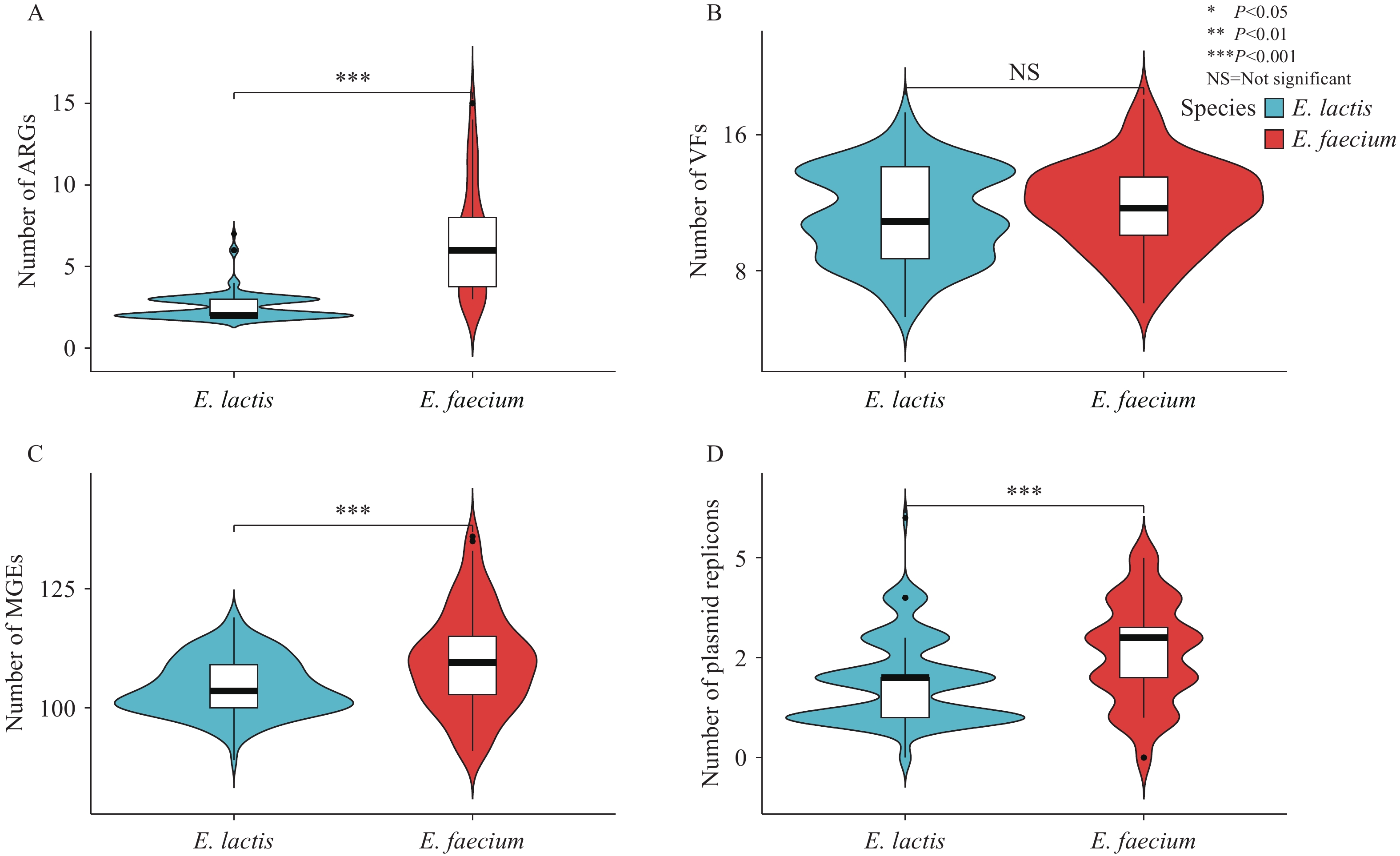 Figure 3.
Figure 3.Comparative analysis of E. faecium and E. lactis genomic content. (A) Antibacterial resistance genes, (B) Virulence factor encoding genes, (C) Mobile genetic elements; (D) Plasmid replicons.
Abbreviation: ARG=antibacterial resistance gene; VF=virulence factor encoding gene; MGE=mobile genetic element; E. faecium=Enterococcus faecium; E. lactis=Enterococcus lactis.Pan-GWAS analysis identified 267 species-specific genes in E. lactis and 222 in E. faecium (Figure 4A). COG functional classification demonstrated that species-specific genes from both enterococcal species were significantly enriched in three major categories: metabolism, cellular processes and signaling, and information storage and processing (Figure 4B).
 Figure 4.
Figure 4.Pan-genome-wide association study of E. faecium and E. lactis. (A) Species-specific differential genes are highlighted in the red box. (B) Relative abundance of differential genes between E. lactis and E. faecium across distinct Clusters of Orthologous Groups functional categories.
Note: For (A), the colored bands on the right side of the phylogenetic tree represent genomic information for Enterococcus isolates. For (B), functional category explanations are provided on the right side of the figure.
Abbreviation: E. faecium=Enterococcus faecium; E. lactis=Enterococcus lactis.
-
The distinct ABR and genomic characteristics of E. lactis and E. faecium pose different food safety risks. E. faecium demonstrated significantly higher resistance to 12 antimicrobials, a substantially elevated MDR rate, and greater abundance of ARGs, MGEs, and plasmid replicons compared to E. lactis. These genetic elements increase the potential for ABR transmission throughout the food chain, with specific gene-phenotype correlations providing mechanistic explanations for the observed resistance patterns. The unexpected correlation between ermT and β-lactam resistance in E. faecium warrants further investigation to elucidate the underlying mechanisms. Although E. lactis exhibited overall lower ABR, it showed notably higher erythromycin resistance than E. faecium. This elevated resistance is likely attributable to the macrolide resistance gene msrC, which is chromosomally encoded and therefore presents limited risk for horizontal gene transfer to other bacterial species (16).
The widespread distribution of adhesion genes likely contributes substantially to colonization capacity and virulence expression of both E. faecium and E. lactis within the food chain. Previous research has demonstrated that sgrA plays a critical role in bacterial adherence during hospital-associated infections by facilitating biofilm formation in E. faecium (17). The high prevalence of sgrA among E. lactis isolates suggests that this species may harbor underappreciated clinical virulence potential, warranting enhanced surveillance of its pathogenic characteristics throughout food production and distribution systems.
Pan-GWAS analysis revealed substantial inter-species variation in gene content between E. faecium and E. lactis, while their functional gene distributions demonstrated remarkable conservation. This pattern suggests that despite divergent evolutionary trajectories involving species-specific gene acquisition and loss events, both species have maintained similar functional capabilities in essential biological processes (3).
Several limitations should be acknowledged. The absence of sampling data from 2020–2022 and the restriction to five PLADs may limit the generalizability of these findings to other time periods and geographic regions. Future surveillance efforts should expand temporal and spatial coverage while implementing species-specific monitoring strategies to comprehensively assess the evolving ABR landscape of enterococci in food production systems.
This research provides valuable insights for addressing antimicrobial resistance risks in the food chain, a central priority within the One Health framework. By characterizing the distinct ABR profiles and genomic features of E. faecium and E. lactis, this study addresses critical knowledge gaps in species-specific risk assessments of enterococci in Chinese food systems. These findings facilitate the integration of food safety, animal health, and public health perspectives, supporting the development of targeted interventions to prevent ABR transmission across food-animal-human interfaces.
-
Approved by the Ethics Committee of the State Key Laboratory of the China National Centre for Food Safety Risk Assessment (CFSA).
HTML
Sample Collection and Enterococcus Detection and Identification
Whole-genome Sequencing (WGS), Genome Assembly, and ANI Analysis
Antimicrobial Susceptibility Testing (AST)
Bioinformatics Analysis
Statistical Analysis
Prevalence of E. faecium and E. lactis in Food Chain
ABR and ARGs of E. faecium and E. lactis
Genomic content of E. faecium and E. lactis
| Citation: |

|