

 Download:
Download:




-
Since the first human infections with highly pathogenic avian influenza (HPAI) H5N6 virus was detected in Sichuan Province of China in April 2014, a total of 38 human cases and 21 deaths due to H5N6 infection have been reported as of August 6, 2021 in the Western Pacific Region (1). However, the newly emergent HPAI H5N6 virus belonging to the genetic clade 2.3.4.4 of H5 virus subtypes has possessed the capability for binding human-origin SA-α, 6 Gal-linked receptor and has demonstrated more transmissibility than H5N1 virus in a ferret model (2), suggesting that this subtype virus may be of high public health risk.
Guangxi Zhuang Autonomous Region in southern China has a history of human infection with avian influenza virus (AIV). H7N9 virus emerged here (3) and HPAI H7N9 virus hit this region in 2017, with 27 infections and 14 deaths (4).
-
We reported 2 cases infected with the H5N6 virus belonging to genetic clade 2.3.4.4b and clade 2.3.4.4h in Guangxi, China in 2021. These 2 cases occurred in two different cities. A case was admitted to the intensive care unit (ICU) due to severe clinical symptoms and was subsequently tested positive for H5N6. The other case was detected from the influenza-like illness (ILI) surveillance system. When a case is found, the local CDC immediately conducts epidemiological and environmental investigations.
On February 16, 2021, a 50-year-old male (Patient A) developed a fever with chest tightness, tightness of breath, headache, cough, sputum, and pneumonia, and was transferred to the Respiratory Department of Hechi People’s Hospital for hospitalization on February 21. Patient A had suffered from rheumatoid arthritis for more than 10 years, and he was found to be H5N6 influenza virus positive on February 26. He had a history of exposure to infected poultry 9 days before the onset of illness and died on March 2.
On July 6, 2021, a 61-year-old female (patient B) developed a fever with a maximum temperature of 38.5 °C and was admitted to the Second People’s Hospital of Guangxi for treatment on July 9. After 6 days, she was confirmed to be positive for H5N6 by Guangxi CDC. She had a history of chronic gastritis and denied having contact with live poultry.
Viral RNA from the throat swabs of the two patients was extracted using the QIAamp® Viral RNA Mini Kit (Qiagen, Germany), according to the manufacturer’s instructions. Specific real-time reverse transcriptase polymerase chain reaction (RT-PCR) assays with specific primer and probe sets for detecting avian influenza A (H5N6). The viral genomes were reverse-transcribed and amplified using Easy-Fast FluA whole genome amplification one-step kit (Xinlihechuang Technology Co., Ltd.). The sequencing libraries were prepared using the Illumina Nextera® XT Library Prep Kit. The final viral-enriched libraries were sequenced using the Illumina MiSeq platform (Illumina, San Diego, USA). The whole genome sequences of the 2 avian influenza A (H5N6) strains were assembled and obtained using CLC Genomics Workbench 9.5.2, and the 2 strains were designated A/GX-hechi/01/2021(H5N6) (GX01) from Patient A and A/GX-guilin/11151/2021(H5N6) (GX11151) from Patient B. The genomes were aligned with the reference genomes downloaded from GISAID (
www.gisaid.org ) using MAFFT v7.037b (https://mafft.cbrc.jp/alignment/software/ ). The genetic and evolutionary analyses were conducted in MEGA 7.0.14 (https://www.megasoftware.net/ ).Through sequencing and splicing, 8 gene fragments of the virus were successfully extracted. Some mutations related to viral replication, receptor-binding, mammalian virulence-related markers and drug-resistance related markers were detected. The PB2 and MP genes of GX01 virus showed high homology with H9N2 viruses, and the following genes showed high homology with H5N6 viruses. The HA and NP genes of GX11151 virus showed high homology with H5N8 and H9N2 viruses, respectively, and the following genes showed high homology with H5N6 viruses (Table 1).
Virus Segment Length (bp) Strain with the highest similarity GISAD ID Similarity (%) GX01 PB2 2348 A/goose/Fujian/3.15_FZHX0011-O/2018 (H9N2) EPI1816470 92.21 PB1 2341 A/Guangxi/31906/2018 (H5N6) EPI1352803 100.00 PA 2229 A/chicken/Miyazaki/2-4C/2017 (H5N6) EPI1891595 96.55 HA 1773 A/chicken/Anhui/8.28_YHZGS017-O/2018 (H5N6) EPI1825343 97.68 NP 1565 A/Guangxi/31906/2018 (A/H5N6) EPI1352798 98.98 NA 1431 A/Env/Guangdong/Qingyuan/C18285099/2018 (H5N6) EPI1366600 98.25 MP 1027 A/chicken/Shanxi/06.28_TGRL001-O/2018 (H9N2) EPI1833450 98.64 NS 875 A/Env/Guangdong/Dongguan/C172863577/2017 (H5N6) EPI1366948 98.86 GX11151 PB2 2342 A/Env/Guangdong/zhanjiang/C17277335/2017 (H5N6) EPI1366759 96.63 PB1 2344 A/Env/Guangdong/zhanjiang/C18277136/2018 (H5N6) EPI1366684 97.78 PA 2233 A/chicken/Miyazaki/2-4C/2017 (H5N6) EPI891595 97.08 HA 1775 A/chicken/Omsk/0112/2020 (H5N8) EPI1813345 99.44 NP 1565 A/duck/Hunan/5.29_YYGK90P3-OC/2018 (H9N2) EPI1835016 98.72 NA 1433 A/Env/Guangdong/Qingyuan/C18285099/2018 (H5N6) EPI1366600 97.77 MP 1027 A/Sichuan/06681/2021 (A/H5N6) EPI1883262 99.51 NS 875 A/Env/Guangdong/Huizhou/C17280804/2017 (A/H5N6) EPI1366935 98.86 Table 1. Similarity analysis of H5N6 virus sequences from the two cases in Guangxi zhuang autonomous region, China, 2021.
Phylogenetic analysis of HA gene was constructed according to the World Health Organization (WHO) reference sequence. Their nucleotide sequence showed a similarity of 90.7% between each other. The HA gene of GX01 falls into clade 2.3.4.4h, while GX11151falls into clade2.3.4.4b (Figure 1).
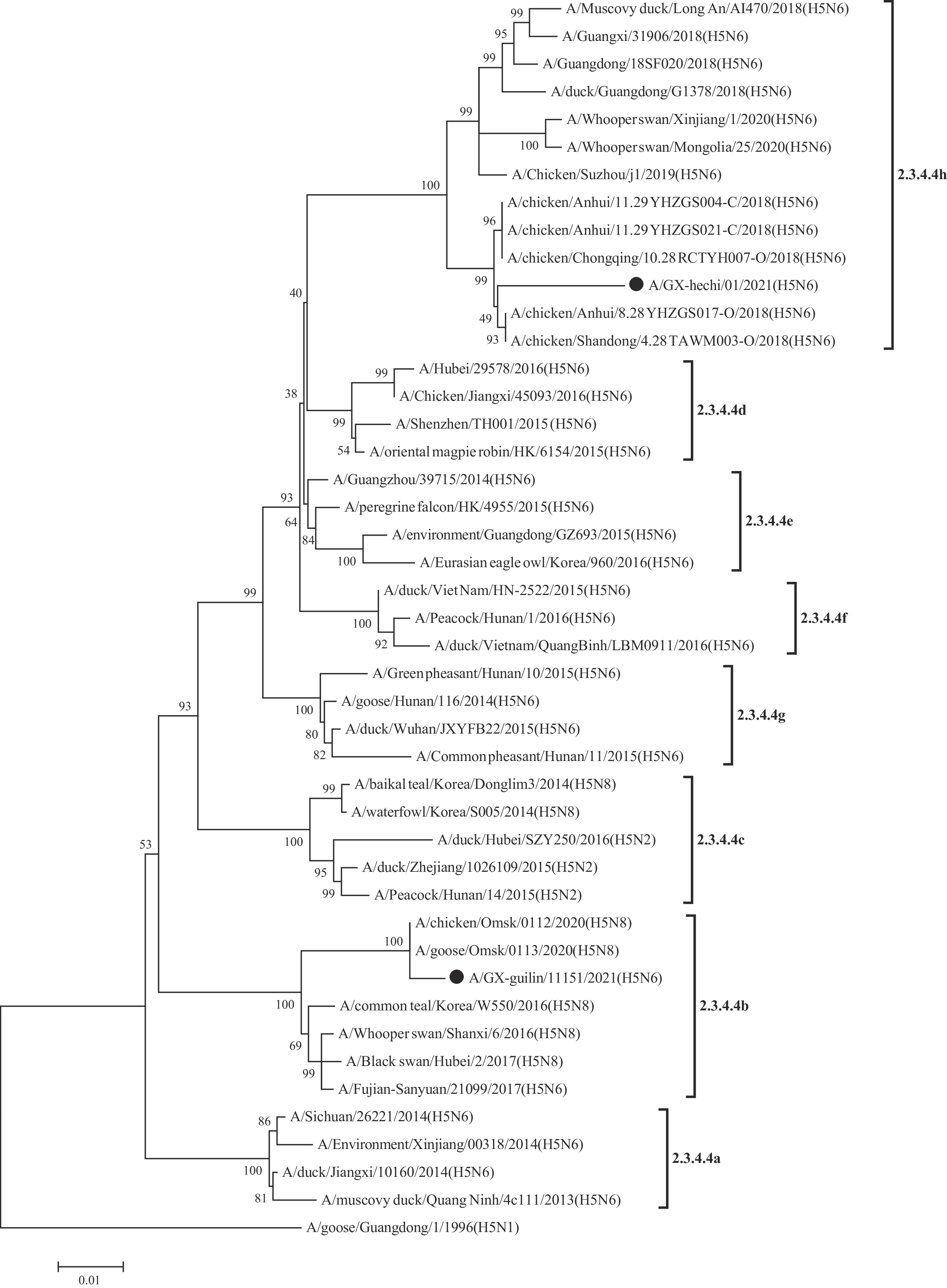 Figure 1.
Figure 1.Phylogenetic relationships of A (H5) clade 2.3.4.4 HA genes using the maximum likelihood method with 1,000 bootstrap.
Note: The two Guangxi strains were indicated by black dots.The HA cleavage site of two viruses possessed a multiple basic amino acids motif, indicating potentially high pathogenicity in chickens. The receptor-binding site at the 222–224 motif was QGG of GX01 virus and QRG of GX11151 virus, respectively, suggesting that these viruses preferred binding to avian-like receptors (α 2,3 SA) (5). However, D94N (6), S133A (7), and T156A (8) mutations in GX01 HA gene, S133A, D155N (8), T156A, and T188I (7) mutations in the GX11151 HA gene increased virus binding to human-like receptors (α2-6 SA). GX01 virus exhibited 8 potential glycosylation sites at 27, 39, 70, 140, 180, 301, 498, and 557 (H5 numbering). GX11151 virus exhibited seven potential glycosylation sites at 27, 39, 180, 209, 301, 498, and 557 (Table 2).
Gene GX01 GX11151 SC26221 GX31906 GZ39715 HB29578 Phenotypic characteristics HA (H5 no.) D94N N S N N N N Increased virus binding to α2-6 SA S133A A A A A A A Increased pseudovirus binding to α2-6 SA D155N D N D D D D Increased virus binding to α2-6 SA T156A A A A A T A T188I T I T A T T Increased pseudovirus binding to α2-6 SA A263T T T T T T T The residue is related to virulence. 222-224 QGG QRG QRG QRG QRG QSG 222–224 QS(R)G avian-like α2–3
receptor binding preferenceCleavage peptides RERRRKR REKRRKR REKRRKR RERRRKR RERRRKR RERRRKR Highly pathogenic avian influenza NA (N6 no.) E119D/V E E E E E E Antiviral oseltamivir resistance A247V A A A A A A H274Y H H H H H H R293K R R R R R R R372K R R R R R R PB2 K389R R R R R R K Enhanced growth capacity in human and mammalian cells V598T/I T T T T T V PB1 I368V I I I I I V Transmissible among ferrets PA N409S S S S S S N Increased virus replicative ability in mammalian systems M1 N30D D D D D D D Increased virulence in mice T139A A A T T T A T215A A A A A A A M2 D21G G D D D D G Antiviral amantadine resistance L26F/I L L L L L L A30T A A A A A A S31N N S S S S N G34E G G G G G G NS1 P42S S S S S S S Increased virulence in mice 80-84Del Yes Yes Yes Yes Yes No L98F M M M M M M Table 2. Molecular features of the genes of H5N6 viruses isolated from humans.
A263T mutation in HA gene was detected in both strains, suggesting that the virulence was enhanced. However, there was no mutation associated with resistance to NA inhibitors in the NA gene of the two strains. K389R, V598T/I mutations of PB2 gene, and N409S mutation of PA gene, which could increase virus replicative ability in mammals, were observed in both strains. Some mutations increasing virulence in mice were detected in our strains, such as N30D, T139A, and T215A of M1 gene and P42S and 80–84 deletion of NS1 gene. The M2 gene of GX01 had the mutations of D21G and S31N, suggesting that the strain was resistant to amantadine, but GX11151 was not observed (Table 2).
-
The previous study showed H5N6 has replaced H5N1 as one of the dominant AV subtypes in southern China (9). The avian influenza A (H5N6) virus continues to threaten human life and health. From January to July 2021, 2 cases were reported in Guangxi zhuang autonomous region. Compared with 4 H5N6 infections reported from 2016 to 2019, and the number of infections cases was slightly higher than that in previous years. It is necessary to sequence and analyze the virus. The case infected with GX01 had obvious clinical symptoms and a history of exposure to dead poultry. Homology analysis showed that GX01 virus was a recombinant virus of H5N6 and H9N2, while GX11151 was a recombinant virus of H5N8, H5N6, and H9N2. Previous study showed H5N6 lineage has been co-circulating in different regions in China (10-11). The mutations in the important sites of proteins of avian influenza virus may change the adaptivity, virulence, tissue tropism, and infectivity. The cleavage between HA1 and HA2 proteins of HA gene of these two viruses were multiple continuous basic amino acids motif (RERRRKR), indicating that they possessed potentially high pathogenicity in chickens. The 222–224 receptor binding site of HA gene suggested that the viruses were avian-like receptors (α 2,3 SA). Studies have shown that the sensitivity of viruses carrying H274Y mutation in NA protein to oseltamivir decreased 1,000 times. In addition, when the NA protein of the virus carries E119A/D/V, A247V, R293K, and R372K mutations, it will cause different degrees of resistance to oseltamivir and zanamivir (12-13). Fortunately, these mutations were not found in our virus. Therefore, using NA inhibitors to treat the 2 cases infected with avian influenza A (H5N6) was still a good choice.
The median age of the two cases was over 50 years old. Like other studies previously reported, the elderly people may be more vulnerable to avian influenza (3). One case was reported through ILI surveillance system (14), indicating that the ILI system was beneficial for the detection of avian influenza cases to a certain extent. However, this may be just the tip of the iceberg, and perhaps many mild cases have not been detected.
This study has certain limitations. Sampling of the environment exposed by the cases, and poultry around the living environment were missing, resulting in lacking of laboratory tests. One reason was that the period from onset to reporting was too long for sampling, in spite of dead poultry found in epidemiology survey, and another reason was that patient B had no clear history of exposure to poultry.
At present, coronavirus disease (COVID-19) has caused a worldwide pandemic (15), and over 200 million people have been infected. Meanwhile, the increasing trend of human infection with avian influenza virus has become an important public health issue that cannot be ignored, alerting us that COVID-19 and avian influenza may be simultaneously prevalent in some regions.
-
Dr. Yu Lan and Dr. William J. Liu.
HTML
| Citation: |

|